 2015-04-30
2015-04-30 2997
2997Сокращение — это изменение механического состояния миофибриллярного аппарата мышечных волокон под влиянием нервных импульсов.
В 1939 г В.А. Энгельгардтом и М.Н. Любимовой было установлено, что миозин обладает свойствами фермента аденозинтрифосфатазы, расщепляющей АТФ. Вскоре было установлено, что при взаимодействии актина с миозином образуется комплекс — актомиозин, ферментативная активность которого почти в 10 раз выше активности миозина (А. С. Уент - Дьорди, 1940). В этот период и начинается разработка современной теории мышечного сокращения, которая получила название теории скользящих нитей. Согласно этой теории «скольжения» в основе сокращения лежит взаимодействие между актиновыми и миозиновыми нитями миофибрилл вследствие образования поперечных мостиков между ними.
Во время скольжения сами актиновые и миозиновые нити не укорачиваются, но длина саркомера изменяется. В расслабленной, а тем более растянутой мышце активные нити располагаются дальше от центра саркомера, и длина саркомера больше. При изотоническом сокращении мышцы актиновые нити скользят по направлению к центру саркомера вдоль миозиновых нитей. Нити актина прикреплены к Z-мембране, тянут ее за собой, и саркомер укорачивается. Суммарное укорочение всех саркомеров вызывает укорочение миофибрилл, и мышца сокращается.
В настоящее время принята следующая модель скольжения нитей актина.
Импульс возбуждения по двигательному нейрону достигает нервно-мышечного синапса — концевой пластинки, где освобождается ацетилхолин, который взаимодействует с постсинаптической мембраной, и в мышечном волокне возникает потенциал действия, т.е. наступает возбуждение мышечного волокна.
При связывании ионов Са++ с тропонином (сферические молекулы которого «сидят» на цепях актина) последний деформируется, толкая тропомиозин в желобки между двумя цепями актина. При этом становится возможным взаимодействие актина с головками миозина и возникает сила сокращения. Головки миозина совершают «гребковые» движения и продвигают актиновую нить по направлению к центру саркомера (рис. 82).
Головок у миозиновых нитей множество, они тянут актиновую нить с объединенной, суммарной силой. При одинаковом гребковом движении головок саркомер укорачивается примерно на 1 % его длины (а при изотоническом сокращении саркомер мышцы может укорачиваться на 50 % длины за десятые доли секунды), следовательно, поперечные мостики должны совершать примерно 50 «гребковых» движений за тот же промежуток времени.
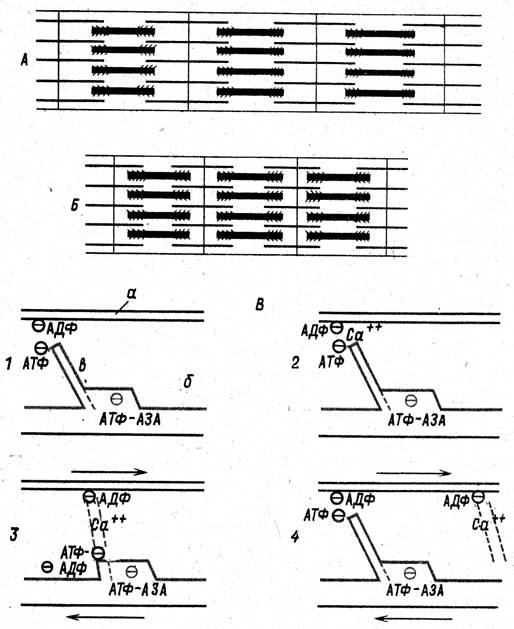
Рис. 82. Схема механизма сокращения миофибриллы:
А- расслабленная и Б- сократившаяся миофибрилла; В- схема последовательности изменений мостика миозиновой нити: а- актиновая нить; б- миозиновая нить; в- мостик; 1- состояние покоя; 2- взаимодействие ионов Са++ с молекулами АДФ и АТФ; 3- укорочение мостика и расщепление АТФ до АДФ; 4- присоединение новой молекулы АТФ, удлинение мостика и готовность к новому циклу. Стрелки показывают движение нитей актина и миозина навстречу друг другу.
Совокупное укорочение последовательно расположенных саркомеров миофибрилл приводит к заметному сокращению мышцы. Одновременно происходит гидролиз АТФ. После окончания пика потенциала действия активируется кальциевый насос (Са — зависимая АТФ-аза) мембраны саркоплазматического ретикулума. За счет энергии, выделяющейся при расщеплении АТФ, кальциевый насос перекачивает ионы Са++ обратно в цистерны саркоплазматического ретикулума, где Са++ связывается белком кальсеквестрином.
Концентрация ионов Са++ в цитоплазме мышц снижается до 10-8 м, а в саркоплазматическом ретикулуме повышается до 10-3 м.
Снижение уровня Са++ в саркоплазме подавляет АТФ-азную активность актомиозина; при этом поперечные мостики миозина отсоединяются от актина. Происходит расслабление, удлинение мышц в результате пассивного движения (без затрат энергии).
Таким образом, сокращение и расслабление мышцы представляет собой серию процессов, развертывающихся в следующей последовательности: нервный импульс --> выделение ацетилхолина пресинаптической мембраной нервно-мышечного синапса --> взаимодействие ацетилхолина с постсинаптической мембраной синапса --> возникновение потенциала действия --> электромеханическое сопряжение (проведение возбуждения по Т-канальцам, высвобождение Са++ и воздействие его на систему тропонин-тропомиозин-актин) --> образование поперечных мостиков и «скольжение» актиновых нитей вдоль миозиновых --> снижение концентрации ионов Са++ вследствие работы кальциевого насоса --> пространственное изменение белков сократительной системы --> расслабление миофибрилл.
После смерти мышцы остаются напряженными, наступает так называемое трупное окоченение, так как поперечные связи между филаментами актина и миозина не могут разорваться из-за отсутствия энергии АТФ и невозможности работы кальциевого насоса.

