 2015-04-30
2015-04-30 1516
1516Биохимия - биологическая химия, наука, изучающая состав организмов, структуру, свойства и локализацию обнаруживаемых в них соединений, пути и закономерности их образования, последовательность и механизмы превращений, а также их биологическая и физиологическая роль. В зависимости от объекта исследования Б. подразделяют на Б. микробов, растений, животных и человека. Это подразделение условно, т.к. в составе различных объектов и в протекающих в них биохимических процессах много общего. Поэтому результаты исследований, проведённых на микробах, растительных или животных тканях и клетках, взаимно дополняют и обогащают друг друга. Тесно связаны между собой и разные направления биохимических исследований, однако принято делить Б. на статическую, занимающуюся преимущественно анализом состава организмов, динамическую, изучающую превращения веществ, и функциональную, выясняющую, какие химические процессы лежат в основе различных проявлений жизнедеятельности. Это последнее направление исследований иногда выделяют под специальным названием физиологическая химия. Вся совокупность химических реакций, протекающих в организмах, включая усвоение веществ, поступающих извне (ассимиляция), и их расщепление (диссимиляция) вплоть до образования конечных продуктов, подлежащих выделению, составляет сущность и содержание обмена веществ — главного и постоянного признака всего живого. Понятно, что изучение обмена веществ во всех деталях — одна из основных задач Б. Биохимические исследования охватывают очень широкий круг вопросов: нет такой отрасли теоретической или прикладной биологии, химии и медицины, которая не была бы связана с Б., поэтому современная Б. объединяет ряд смежных научных дисциплин, ставших с середины 20 в. самостоятельными. Накопление биохимических сведений и формирование Б. в 16—19 вв. Б. сформировалась как самостоятельная наука в конце 19 в., хотя истоки её относятся к далёкому прошлому. С 1-й половины 16 в. и до 2-й половины 17 в. свой вклад в развитие химии и медицины вносили ятрохимики (химики-врачи): немецкий врач и естествоиспытатель Ф. Парацельс, голландские учёные Я. Б. ван Гельмонт, Ф. Сильвий и др., занимавшиеся исследованием пищеварительных соков, жёлчи, а также процессов брожения (см. Ятрохимия). Сильвий, прославленный врач, придавал особенно большое значение правильному соотношению в организме человека кислот и щелочей; он полагал, что в основе многих, если не всех, болезней лежит расстройство этого соотношения. Большая часть принятых ятрохимиками положений была наивной, полной заблуждений; однако нельзя забывать, что научной химии тогда ещё не существовало. Наиболее общей теорией, господствовавшей в науке того времени, была теория т. н. флогистона. Тем не менее, балансовые опыты на человеке с точным учётом массы тела и выделений были проведены итальянским учёным С. Санторио в начале 17 в. Эти опыты привели к описанию "perspiratio insensibilis" — потери массы за счёт "нечувствуемого пропотевания". Великие открытия в области физики и химии 18 и начала 19 вв. (открытие ряда простых веществ и соединений, формулировка газовых законов, открытие законов сохранения материи и энергии) заложили научный фундамент общей химии. После открытия в составе воздуха кислорода голландский ботаник Я. Ингенхауз смог описать постоянное образование растением СО2 и выделение на солнечном свету зелёными частями растения кислорода. Опытами Ингенхауза было положено начало исследованию дыхания растений и процессов фотосинтеза, детальное изучение которых продолжается и в настоящее время. В конце 1-й четверти 19 в. было известно очень ограниченное количество органических веществ. В учебнике немецкого химика Л. Гмелина, изданном в 1822, упоминается лишь 80 органических соединений. Задачи и возможности органической химии в то время оставались неясными. Шведский учёный И. Берцелиус считал, что органические тела разделяются на два четко разграниченных класса — на растения и животные; сущность живого тела основана не на его неорганических элементах, а на чём-то ином. Это нечто, что он называет "жизненной силой", лежит целиком за пределами неорганических элементов. Берцелиус выражает сомнение в том, что люди когда-либо сумеют искусственно производить органические вещества и подтвердить анализ синтезом (1827). Несостоятельность таких типичных для витализма позиций выявилась очень скоро. Уже в 1828 немецкий химик Ф. Вёлер, ученик Берцелиуса, получил синтетическим путём мочевину, описанную в 18 в. французским учёным Г. Руэлем в качестве составной части мочи млекопитающих. Вскоре последовали синтезы других, как природных органических соединений, так и искусственных, неизвестных в природе, т.е. рушилась стена, отделявшая органические соединения от неорганических. Начиная со 2-й половины 19 в. органическая химия становится всё больше химией синтетической, усилия которой направляются на получение новых соединений углерода, особенно имеющих промышленное значение; в её задачи уже не входит исследование состава растительных и животных объектов. Эти сведения поступали случайно в результате побочной работы химиков, ботаников, физиологов растений и животных, а также патологов и врачей, включавших в круг своих интересов химические исследования. Так, в 1814 русский химик К. С. Кирхгоф описал осахаривание крахмала под влиянием вытяжки из проросших семян ячменя: действие амилазы. К середине 19 в. были описаны и другие ферменты: амилаза слюны, расщепляющая полисахариды; пепсин желудочного сока и трипсин сока поджелудочной железы, расщепляющие белки. Берцелиус ввёл в химию понятие катализаторов, к числу которых были отнесены все известные в то время ферменты. В 1835 французский химик М. Шеврёль описал в составе мышц креатин, несколько позднее в моче был найден близкий к нему по структуре креатинин. Содержание в скелетных мышцах молочной кислоты и её накопление при работе установил немецкий химик Ю. Либих. В 1839 он же выяснил, что в состав пищи входят белки, жиры и углеводы, являющиеся главными составными частями животных и растительных организмов. В середине 19 в. была установлена структура жира и осуществлен его синтез французским химиком П. Бертло; синтез углеводов был проведён русским учёным А. М. Бутлеровым; он же предложил теорию строения органических соединений, сохранившую своё значение и поныне. Систематическое исследование белков было начато голландским врачом и химиком Г. И. Мульдером в 30-е гг. 19 в. и интенсивно продолжалось многими авторами во все последующие годы. В то же время в связи с описанием дрожжевых клеток (К. Коньяр-Латур во Франции и Т. Шванн в Германии, 1836—38) активно начали изучать процесс сбраживания сахара и образования спирта, издавна привлекавший к себе внимание. В числе учёных, изучавших брожение, были Ю. Либих и французский учёный Л. Пастер. Пастер пришёл к выводу, что брожение — биологический процесс, в котором обязательно участвуют живые дрожжевые клетки. Либих же рассматривал сбраживание сахара как сложную химическую реакцию. В этот спор была внесена ясность, когда русский химик М. М. Манассеина (1871) и особенно четко немецкий учёный Э. Бухнер (1897) доказали способность бесклеточного дрожжевого сока вызывать алкогольное брожение. Т. о. была подтверждена принципиальная правильность химической теории действия ферментов, которую Либих сформулировал в 1870; основные принципы этой теории сохранили своё значение и теперь. Постепенно количество накопившихся сведений относительно химического состава растительных и животных организмов и протекающих в них химических реакций стало значительным, в связи с чем были осуществлены попытки их систематизации и объединения в учебных руководствах. Наиболее ранние из них — учебники И. Зимона (1842) и Либиха (1847), изданные в Германии, и учебник физиологической химии А. И. Ходнева, вышедший в России (1847).
Возникновение и развитие современных направлений биохимии. В конце 19 века и в 20 в. развитие Б. приобрело выраженный специализированный характер в зависимости от разрабатываемой проблемы и объекта исследования. Б. растений развивалась по преимуществу на кафедрах ботаники и физиологии растений. Тесно связана с ней и Б. микроорганизмов. Белки, углеводы, липиды, витамины, являющиеся составными частями растений, животных и микроорганизмов, исследовали биохимики всех стран на самых различных объектах. Характерными для растений и микроорганизмов можно считать гликозиды, дубильные вещества, эфирные масла, алкалоиды, антибиотики и др. т. н. вещества вторичного происхождения. Из перечисленных соединений ряд гликозидов был синтезирован при участии ферментов французским химиком Э. Буркло и его сотрудниками (1911—18). В расшифровке строения антоцианов — гликозидов, входящих в состав пигментов цветов и плодов, — исключительную роль сыграли классические работы немецкого химика Р. Вильштеттера (1910—15). Группа алкалоидов (азотистых гетероциклических веществ основного характера) изучалась немецким химиком А. Гофманом (1890—1900). Позднее алкалоиды изучали выдающиеся исследователи (Р. Вильштеттер, Л. Пикте — Швейцария; русские химики А. П. Орехов, А. А. Шмук и многие др.). Эфирные масла, терпены успешно исследовали также крупные представители химии и биохимии: Перкин младший (Великобритания), Г. Эйлер (Швеция) и др. Выдающуюся роль в развитии Б. растений в России (конец 19 в. — 1-я половина 20 в.) сыграли профессор Петербургского университета А. С. Фаминцын, его ученики Д. И. Ивановский, открывший вирусы, и И. П. Бородин, изучавший окислительные процессы в организме растений и их связь с превращениями белков. Работы С. П. Костычева (профессор Петербургского университета, позднее — ЛГУ) по анаэробному обмену углеводов и дыханию у растений обогатили химическую физиологию открытием новых промежуточных продуктов брожения, формулировкой оригинальных взглядов на сущность окислительных процессов, на обмен белков и фиксацию азота растениями. Много сделал профессор Варшавского университета М. С. Цвет, разработавший метод хроматографии на колонках, используемый и в настоящее время. Московская школа физиологов и биохимиков растений была представлена К. А. Тимирязевым, исследовавшим фотосинтез и химию хлорофилла. Его ученики — В. И. Палладин, разрабатывавший проблему биологического окисления, Д. П. Прянишников, изучавший азотистый обмен растений, В. С. Буткевич, обогативший теоретическую Б. исследованиями белков и белкового обмена растений, А. Р. Кизель, изучавший обмен аргинина и мочевины у растений и структурные элементы протоплазмы клеток, — явились создателями крупных школ и оригинальных направлений современной общей и эволюционной Б., а также физиологии и Б. растений, плодотворно развивающихся и в 3-й четверти 20 в. представители Б. микроорганизмов и Б. растений решали много общих задач, связанных с изучением природных соединений (в т. ч. и высокомолекулярных), их структуры, путей образования и расщепления, характеристики ферментов, участвующих в этих процессах. Следует отметить, что микроорганизмы постепенно стали излюбленным объектом для различных энзимологических исследований и для разработки проблем биохимической генетики. Все эти исследования создали прочную базу для разработки многих частных проблем, в том числе и промышленной Б. К ним относятся получение новых антибиотиков, разработка методов их очистки, поиски условий, благоприятных для микробиологического синтеза не только антибиотиков, но и других биологически активных соединений — витаминов, дефицитных аминокислот, нуклеотидов и т.д. Б. животных и человека (медицинская и физиологическая химия). Большое значение для развития этой ветви Б. имели многочисленные школы физиологов, химиков, патологов и врачей, работавших в разных странах. Во Франции в лаборатории физиолога К. Бернара в составе печени млекопитающих был открыт гликоген (1857), изучены пути его образования и механизмы, регулирующие его расщепление; здесь же Л. Корвизар (1856) открыл в поджелудочном соке фермент трипсин. В Германии в лабораториях Ф. Хоппе-Зейлера, А. Косселя, Э. Фишера, Э. Абдергальдена, О. Хаммарстена и др. подробно изучались простые и сложные белки, их структура и свойства, вещества, образующиеся при искусственном их расщеплении путём нагревания с кислотами и щёлочами, а также под влиянием ферментов. В Англии Ф. Хопкинс, основатель школы биохимиков в Кембридже, занимался исследованием аминокислотного состава белков, открыл триптофан, глутатион, изучал роль аминокислот и витаминов в питании. Существенный вклад в развитие Б. в конце 19 — начале 20 вв. внесли русские учёные, работавшие на кафедрах высших учебных заведений и в специализированных институтах. В Военно-медицинской академии А.Я. Данилевский и его сотрудники разрабатывали проблемы химии белка, методы выделения и очистки ферментов, изучали механизм их действия и условия обратимости ферментативных реакций. В Институте экспериментальной медицины М. В. Ненцкий исследовал химию порфиринов, биосинтез мочевины, а также ферменты бактерий, вызывающие разложение аминокислот. Особенно плодотворным было содружество лабораторий А. Я. Данилевского и М. В. Ненцкого с лабораторией И. П. Павлова при исследовании пищеварения и образования мочевины в печени. В Московском университете В. С. Гулевич подробно и успешно исследовал азотистые экстрактивные (небелковые) вещества мышц и открыл ряд новых соединений оригинальной структуры (карнозин, карнитин и др.). Предметом многочисленных исследований было и остаётся подробное изучение разнообразных ферментативных реакций, протекающих в паренхиматозных органах, главным образом в печени, и обусловливающих нормальное течение процессов обмена веществ. Большое внимание во 2-й половине 19 и в 20 вв. было уделено биохимическому исследованию возбудимых тканей, главным образом мозга и мышц. В СССР разработка этих проблем осуществлялась А. В. Палладиным, Г. Е. Владимировым, Е. М. Крепсом, их учениками и сотрудниками. К середине 20 в. нейрохимия представляла одно из сформировавшихся самостоятельных направлений. Подверглась всестороннему изучению Б. крови. Дыхательная функция крови (т. е. связывание и отдача кровью углекислого газа и кислорода), изучавшаяся в середине 19 в. в лаборатории К. Людвига в Вене, подробно исследовалась в дальнейшем в разных странах. Полученные данные привели к анализу структуры и свойств гемоглобина в норме и патологии, к детальному изучению реакции между гемоглобином и кислородом и выяснению закономерностей кислотно-щелочного равновесия. Крупных успехов Б. достигла в изучении витаминов, гормонов, минеральных веществ, в частности микроэлементов, их распространения в различных организмах, физиологической роли, механизма действия и регулирующих влияний на ферментативные реакции и процессы обмена веществ. Большое значение имеет проблема связи структуры и функции, которая характеризует также задачи биохимической фармакологии, когда речь идёт о лекарственных средствах и исследовании первичного механизма их действия, осуществляемого вмешательством в ферментативные реакции, составляющие основу процессов обмена веществ. В середине 20 в. самостоятельное значение приобрели биохимические исследования, проводившиеся в клиниках и посвященные изучению биохимических особенностей организма, химического состава крови, мочи и других жидкостей и тканей больного человека. Это направление, получившее широкое развитие, составляет основное содержание клинической биохимии. Витаминология. В лаборатории Г. А. Бунге молодой русский врач Н. И. Лунин первый описал в 1880 в составе молока добавочные факторы питания. В 1896 аналогичное наблюдение было сделано голландским врачом К. Эйкманом, описавшим присутствие важного для организма фактора в рисовых отрубях. Польский исследователь К. Функ в 1912 выделил активное начало в кристаллическом виде и назвал его витамином. Работы этого направления получили широкое развитие; постепенно были открыты многие другие витамины, и сейчас витаминология представляет один из весьма важных разделов Б., а также науки о питании. Б. гормонов. Работы, связанные с анализом химической структуры продуктов жизнедеятельности желёз внутренней секреции — гормонов, путей их образования в организме, механизма действия и возможного осуществления лабораторного синтеза, представляют одно из важных направлений биохимических исследований. Б. стероидных гормонов — часть общей проблемы Б. стеринов. Достигнутые в этой области успехи в значительной мере связаны с использованием меченных по углероду (С14) исходных и промежуточных соединений. Самая тесная связь установилась между широким фронтом исследований белковых веществ и специальным изучением структуры и функций гормонов белковой природы. Изучение гормональной активности тех или других препаратов невозможно без глубокого анализа биохимического механизма их действия. Т. о., данные по химии и Б. гормонов в равной мере обогащают эндокринологию и Б. Энзимология — учение о ферментах, вполне самостоятельная область Б. В ней проблема строения белков-ферментов тесно переплетается с физико-химическими проблемами — химической кинетикой и катализом. В 3-й четверти 20 в. внесено много нового в представления о структуре ферментов, о их присутствии в нативном состоянии в виде сложных комплексов. Анализ строения ферментов в сопоставлении с проявляемой ими в разных условиях активностью позволил выяснить значение отдельных аминокислот (главным образом цистеина, лизина, гистидина, тирозина, серина и т.д.) в формировании активного центра ферментов. Выяснены структура многих коферментов, их значение для ферментативной активности, а также связь между коферментами и витаминами. Большой вклад в развитие энзимологии в первой половине 20 в. внесли Р. Вильштеттер, Л. Михаэлис, Г. Эмбден, О. Мейергоф (Германия), Дж. Самнер, Дж. Нортроп (США), Г. Эйлер (Швеция), А. Н. Бах (СССР). Много сделали продолжающие активно работать создатели крупных школ и направлений: О. Варбург (Западный Берлин), Ф. Линен (ФРГ), Р. Питерс, Х. Кребс (Великобритания), Х. Теорелль (Швеция), Ф. Линман, Д. Кошленд (США), А. Росси-Фанелли (Италия), Ф. Шорм (Чехословакия), Ф. Штрауб (Венгрия), Т. Барановский, Ю. Хеллер (Польша) и многие другие. В СССР эту область исследований представляют: В. А. Энгельгардт и М. Н. Любимова, установившие ферментативную активность мышечных белков, в частности аденозинтрифосфатазную активность миозина и процесс окислительного фосфорилирования; А.Е. Браунштейн, открывший совместно с М. Г. Крицман процесс переноса аминогруппы (переаминирование); А. И. Опарин и Л.Л. Курсанов, изучавшие роль структуры клеток в проявлении активности ферментов; С.Р. Мардашев, успешно исследовавший декарбоксилирование аминокислот, и др. Исследования сложных комплексов ферментов проводятся в лабораториях Л. Рида (США), М. Койке (Япония), В. Санади (США), Ф. Линена (ФРГ), С. Е. Северина (СССР) и др. Советский учёный В. А. Белицер значительно углубил представления об энергетической эффективности открытого В. А. Энгельгардтом дыхательного пути образования богатых энергией соединений; Г. Е. Владимиров уточнил количество энергии (10 кал, или 42 дж), освобождающееся при гидролизе АТФ (см. Аденозинфосфорные кислоты). Работы этого направления, сначала остававшиеся единичными, в 50-е и последующие годы получили очень широкое развитие главным образом в результате исследований Д. Грина и Б. Чанса, А. Ленинджера, Э. Рэккера (США), Э. Слатера (Нидерланды), Л. Эрнстера (Швеция) и др. В СССР эта проблема разрабатывалась в МГУ и ЛГУ на кафедрах Б., а также в отдельных лабораториях (С. А. Нейфах, В. П. Скулачев и др.). Современные исследования показали также наличие выраженного влияния солевого состава среды и отдельных ионов на ферментативные процессы и важную роль микроэлементов в реализации ферментативной активности. Эволюционная и сравнительная Б. Исследования по Б. животных, растений и микроорганизмов показали, что, несмотря на общность основных биохимических структур и процессов у всех живых организмов, имеются и специфические различия, зависящие от уровня онто- и филогенетического развития изучавшихся объектов. Накопленные факты позволили заложить фундамент сравнительной Б., задача которой — найти закономерности биохимической эволюции организмов. Большое теоретическое значение имеет проблема происхождения жизни на Земле. Некоторые важные положения теории А. И. Опарина о происхождении жизни получили экспериментальное подтверждение в работах института им. Баха, кафедры Б. растений МГУ и ряда зарубежных лабораторий (И. Оро, С. У. Фокс в США; и др.). Гистохимия. Цитохимия. По мере развития техники морфологических исследований, особенно после введения в практику лабораторной работы электронной микроскопии, открывшей многочисленные, ранее неизвестные структуры в составе клеточного ядра и протоплазмы, перед Б. встали новые задачи. На стыке морфологических и биохимических исследований возникли новые отрасли — гистохимия и цитохимия, изучающие локализацию и превращение веществ в клетках и тканях и использующие биохимические и морфологические методы. Биоорганическая химия. Подробные исследования структуры биополимеров — простых и сложных белков, нуклеиновых кислот, полисахаридов и липидов, а также анализ действия биологически активных низкомолекулярных природных соединений (коферментов, нуклеотидов, витаминов и т.д.) привели к необходимости изучения связи между строением вещества и его биологической функцией. Постановка этого вопроса вызвала развитие исследований, находящихся на грани биологической и органической химии. Данное направление исследований получило наименование биоорганической химии. Молекулярная биология. Разработка методов разделения субклеточных структур (ультрацентрифугирование) и получение отдельно фракций, содержащих клеточные ядра, митохондрии, рибосомы и т.п., позволили детально исследовать состав и биологические функции выделенных образований. Применение методов электрофореза в сочетании с хроматографией дало возможность детально характеризовать высокомолекулярные соединения. Параллельно улучшалась техника аналитических определений, позволявшая исследовать ничтожное количество материала. Это было связано с внедрением в биологию, в том числе и в Б., физических (главным образом оптических) методов исследования (флуорометрия, спектрофотометрия в различных областях спектра, масс-спектрометрия, ядерномагнитный и электронно-парамагнитный резонанс, газово-жидкостная хроматография), с применением радиоактивных изотопов, чувствительных автоматических анализаторов аминокислот, пептидов, нуклеотидов, полярографии, высоковольтного электрофореза и т.д. Всё это привело к появлению ещё одного самостоятельного ответвления Б., тесно связанного с биофизикой и физической химией и названного молекулярной биологией. Составной частью молекулярной биологии можно считать молекулярную генетику, несмотря на некоторые специфические её задачи. Так, например, анализ механизма возникновения ряда наследственных нарушений обмена веществ и функций организма позволил выяснить роль выпадения или извращения биосинтеза тех или иных белковых веществ, обладающих ферментативной, иммунной или другой биологической активностью. Сюда относятся также исследования нарушений в обмене углеводов, аминокислот (например, фенилаланина, тирозина, триптофана и др.), образования патологических форм гемоглобина и т.д. Благодаря развитию новых методов исследования Б. в 1950—1970 гг. достигла крупных успехов. Это, прежде всего — выяснение строения белков, определение последовательности расположения в них аминокислот. Впервые была выяснена последовательность расположения аминокислот в гормоне белковой природы — инсулине — английским биохимиком Ф. Сангером, затем в ферменте рибонуклеазе К.Хёрсом, С. Муром и У.Стейном (США), разработавшими метод автоматического анализа аминокислот, вошедший в практику биохимических лабораторий. Тот же фермент — рибонуклеазу, полученную из разных источников, изучали К. Анфинсен (США), Ф. Эгами (Япония) и др. Последовательность расположения аминокислот в ряде протеолитических ферментов установили Ф. Шорм и Б. Кейль с сотрудниками (Чехословакия), Б. Хартли (Великобритания) и др. Большое достижение Б. 60-х гг. 20 в. — химический синтез гормонов — адренокортикотропного гормона, молекула которого содержит 23 аминокислоты (в природном гормоне 39 аминокислот), и инсулина, молекула которого состоит из 51 аминокислоты, фермента рибонуклеазы (124 аминокислоты). С большим успехом использовали английские учёные М. Перуц, Дж. Кендрю и их сотрудники рентгеноструктурный анализ для выяснения строения миоглобина и гемоглобина. В 1956—67 полностью была определена структура лизоцима английским биохимиком Д. Филлипсом и др. Не менее значительны успехи, достигнутые в анализе сложных белков, нуклеопротеидов, нуклеиновых кислот и нуклеотидов. Триумфом Б., молекулярной биологии и генетики явились исследования, показавшие роль нуклеиновых кислот в биосинтезе белков и установившие предопределяющее влияние нуклеиновых кислот на строение и свойства синтезируемых в клетке белков. Этими работами были выяснены биохимические основы передачи признаков по наследству от поколения к поколению. Трудно переоценить также значение исследований, определивших последовательность нуклеотидов в составе транспортных рибонуклеиновых кислот и разработку методов органического синтеза полинуклеотидов. Особенно плодотворно в названных областях работают И. Бьюкенен, Э. Чаргафф, И. Дэвидсон, Д. Дейвис, А. Корнберг, С. Очоа, Дж. Уотсон, М. Уилкинс и др. (США), Ф. Крик, Ф. Сангер (Великобритания).
I.1. Строение клетки. Клетка — элементарная единица строения и жизнедеятельности всех живых организмов (кроме вирусов, о которых нередко говорят как о неклеточных формах жизни), обладающая собственным обменом веществ, способная к самостоятельному существованию, самовоспроизведению и развитию. История открытия. Клеточная теория. Первым человеком, увидевшим клетки, был английский учёный Роберт Гук пробковое дерево так хорошо плавает, Гук стал рассматривать тонкие срезы пробки с помощью усовершенствованного им микроскопа. Он обнаружил, что пробка разделена на множество крошечных ячеек, напомнивших ему монастырские кельи, и он назвал эти ячейки клетками (по-английски cell означает «келья, ячейка, клетка»). В 1675 году итальянский врач М. Мальпиги, а в 1682 году — английский ботаник Н. Грю подтвердили клеточное строение растений. О клетке стали говорить как о «пузырьке, наполненном питательным соком». В 1674 году голландский мастер Антоний ван Левенгук (Anton van Leeuwenhoek, 1632—1723) с помощью микроскопа впервые увидел в капле воды «зверьков» — движущиеся живые организмы (инфузории, амёбы, бактерии). Также Левенгук впервые наблюдал животные клетки — эритроциты и сперматозоиды. Таким образом, уже к началу XVIII века учёные знали, что под большим увеличением растения имеют ячеистое строение, и видели некоторые организмы, которые позже получили название одноклеточных. В 1802—1808 годах французский исследователь Шарль-Франсуа Мирбель установил, что все растения состоят из тканей, образованных клетками. Ж. Б. Ламарк в 1809 году распространил идею Мирбеля о клеточном строении и на животные организмы. В 1825 году чешский учёный Я. Пуркине открыл ядро яйцеклетки птиц, а в 1839 ввёл термин «протоплазма». В 1831 году английский ботаник Р. Броун впервые описал ядро растительной клетки, а в 1833 году установил, что ядро является обязательным органоидом клетки растения. С тех пор главным в организации клеток считается не мембрана, а содержимое. Клеточная теория строения организмов была сформирована в 1839 году немецким зоологом Т. Шванном и М. Шлейденом и включала в себя три положения. В 1858 году Рудольф Вирхов дополнил её ещё одним положением, однако в его идеях присутствовал ряд ошибок: так, он предполагал, что клетки слабо связаны друг с другом и существуют каждая «сама по себе». Лишь позднее удалось доказать целостность клеточной системы. В 1878 году русским учёным И. Д. Чистяковым открыт митоз в растительных клетках; в 1878 году В. Флемминг и П. И. Перемежко обнаруживают митоз у животных. В 1882 году В. Флемминг наблюдает мейоз у животных клеток, а в 1888 году Э. Страсбургер — у растительных.
Строение клетки. Все клеточные формы жизни на Земле можно разделить на два надцарства на основании строения составляющих их клеток: прокариоты (доядерные) — более простые по строению и возникли в процессе эволюции раньше; эукариоты (ядерные) — более сложные, возникли позже. Клетки, составляющие тело человека, являются эукариотическими. Несмотря на многообразие форм, организация клеток всех живых организмов подчинена единым структурным принципам. Содержимое клетки отделено от окружающей среды плазматической мембраной, или плазмалеммой. Внутри клетка заполнена цитоплазмой, в которой расположены различные органоиды и клеточные включения, а также генетический материал в виде молекулы ДНК. Каждый из органоидов клетки выполняет свою особую функцию, а в совокупности все они определяют жизнедеятельность клетки в целом. Прокариотическая клетка (рис 1)
Рис.1
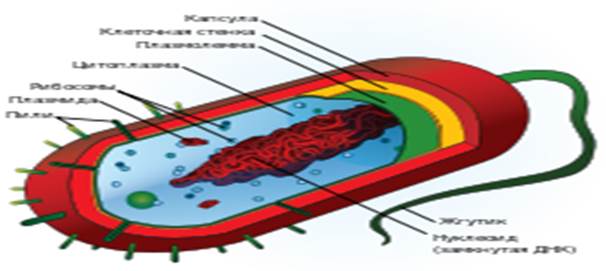
Строение типичной клетки прокариот: капсула, клеточная стенка, плазмолемма, цитоплазма, рибосомы, плазмина, пили, жгутик, нуклеоид. Прокариоты (от лат. pro — перед, до и греч. κάρῠον — ядро, орех) — организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов — линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток — митохондрии и пластиды. Основное содержимое клетки, заполняющее весь её объём, — вязкая зернистая цитоплазма. Эукариоты (эвкариоты) (от греч. ευ — хорошо, полностью и κάρῠον — ядро, орех) — организмы, обладающие, в отличие от прокариот, оформленным клеточным ядром, отграниченным от цитоплазмы ядерной оболочкой. Генетический материал заключён в нескольких линейных двухцепочных молекулах ДНК (в зависимости от вида организмов их число на ядро может колебаться от двух до нескольких сотен), прикреплённых изнутри к мембране клеточного ядра и образующих у подавляющего большинства (кроме динофлагеллят) комплекс с белками-гистонами, называемый хроматином. В клетках эукариот имеется система внутренних мембран, образующих, помимо ядра, ряд других органоидов (эндоплазматическая сеть, аппарат Гольджи и др.). Кроме того, у подавляющего большинства имеются постоянные внутриклеточные симбионты-прокариоты — митохондрии, а у водорослей и растений — также ипластиды. Строение эукариотической клетки (рис 2.) Рис.2
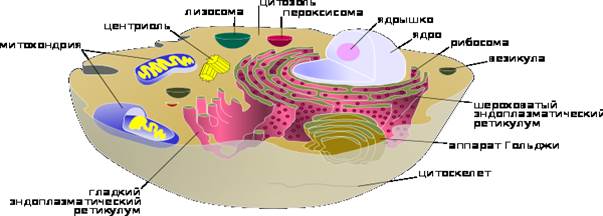
Поверхностный комплекс животной клетки состоит из гликокаликса, плазмалеммы и расположенного под ней кортикального слоя цитоплазмы. Плазматическая мембрана называется также плазмалеммой, наружной клеточной мембраной. Это биологическая мембрана, толщиной около 10 нанометров. Обеспечивает в первую очередь разграничительную функцию по отношению к внешней для клетки среде. Кроме этого она выполняет транспортную функцию. На сохранение целостности своей мембраны клетка не тратит энергии: молекулы удерживаются по тому же принципу, по которому удерживаются вместе молекулы жира — гидрофобным частям молекул термодинамически выгоднее располагаться в непосредственной близости друг к другу. Гликокаликс представляет собой «заякоренные» в плазмалемме молекулы олигосахаридов, полисахаридов, гликопротеинов и гликолипидов. Гликокаликс выполняет рецепторную и маркерную функции. Плазматическая мембрана животных клеток в основном состоит из фосфолипидов и липопротеидов с вкрапленными в неё молекулами белков, в частности, поверхностных антигенов и рецепторов. В кортикальном (прилегающем к плазматической мембране) слое цитоплазмы находятся специфические элементы цитоскелета — упорядоченные определённым образом актиновые микрофиламенты. Основной и самой важной функцией кортикального слоя (кортекса) являются псевдоподиальные реакции: выбрасывание, прикрепление и сокращение псевдоподий. При этом микрофиламенты перестраиваются, удлиняются или укорачиваются. От структуры цитоскелета кортикального слоя зависит также форма клетки (например, наличие микроворсинок).
Структура цитоплазмы. Жидкую составляющую цитоплазмы также называют цитозолем. Под световым микроскопом казалось, что клетка заполнена чем-то вроде жидкой плазмы или золя, в котором «плавают» ядро и другие органоиды. На самом деле это не так. Внутреннее пространство эукариотической клетки строго упорядочено. Передвижение органоидов координируется при помощи специализированных транспортных систем, так называемых микротрубочек, служащих внутриклеточными «дорогами» и специальных белков динеинов и кинезинов, играющих роль «двигателей». Отдельные белковые молекулы также не диффундируют свободно по всему внутриклеточному пространству, а направляются в необходимые компартменты при помощи специальных сигналов на их поверхности, узнаваемых транспортными системами клетки.
Эндоплазматический ретикулум. В эукариотической клетке существует система переходящих друг в друга мембранных отсеков (трубок и цистерн), которая называется эндоплазматическим ретикулумом (или эндоплазматическая сеть, ЭПР или ЭПС). Ту часть ЭПР, к мембранам которого прикреплены рибосомы, относят к гранулярному (или шероховатому) эндоплазматическому ретикулуму, на его мембранах происходит синтез белков. Те компартменты, на стенках которых нет рибосом, относят к агранулярному (или гладкому) ЭПР, принимающему участие в синтезе липидов. Внутренние пространства гладкого и гранулярного ЭПР не изолированы, а переходят друг в друга и сообщаются с просветом ядерной оболочки.
Аппарат Гольджи. Он представляет собой стопку плоских мембранных цистерн, несколько расширенных ближе к краям. В цистернах аппарата Гольджи созревают некоторые белки, синтезированные на мембранах гранулярного ЭПР и предназначенные для секреции или образования лизосом. Аппарат Гольджи асимметричен — цистерны располагающиеся ближе к ядру клетки (цис-Гольджи) содержат наименее зрелые белки, к этим цистернам непрерывно присоединяются мембранные пузырьки — везикулы, отпочковывающиеся от эндоплазматического ретикулума. По-видимому, при помощи таких же пузырьков происходит дальнейшее перемещение созревающих белков от одной цистерны к другой. В конце концов, от противоположного конца органеллы (транс-Гольджи) отпочковываются пузырьки, содержащие полностью зрелые белки.
Ядро. Клеточное ядро содержит молекулы ДНК, на которых записана генетическая информация организма. В ядре происходит репликация — удвоение молекул ДНК, а также транскрипция — синтез молекул РНК на матрице ДНК. В ядре же синтезированные молекулы РНК претерпевают некоторые модификации (например, в процессе сплайсинга из молекул матричной РНК исключаются незначащие, бессмысленные участки), после чего выходят в цитоплазму. Сборка рибосом также происходит в ядре, в специальных образованиях, называемых ядрышками. Компартмент для ядра — кариотека — образован за счёт расширения и слияния друг с другом цистерн эндоплазматической сети таким образом, что у ядра образовались двойные стенки за счёт окружающих его узких компартментов ядерной оболочки. Полость ядерной оболочки называется люменом или перинуклеарным пространством. Внутренняя поверхность ядерной оболочки подстилается ядерной ламиной, жесткой белковой структурой, образованной белками-ламинами, к которой прикреплены нити хромосомной ДНК. В некоторых местах внутренняя и внешняя мембраны ядерной оболочки сливаются и образуют так называемые ядерные поры, через которые происходит материальный обмен между ядром и цитоплазмой.
Лизосома — небольшое тельце, ограниченное от цитоплазмы одинарной мембраной. В ней находятся литические ферменты, способные расщепить все биополимеры. Основная функция — аутолиз — то есть расщепление отдельных органоидов, участков цитоплазмы клетки.
Цитоскелет. К элементам цитоскелета относят белковые фибриллярные структуры, расположенные в цитоплазме клетки: микротрубочки, актиновые и промежуточные филаменты. Микротрубочки принимают участие в транспорте органелл, входят в состав жгутиков, из микротрубочек строится митотическое веретено деления. Актиновые филаменты необходимы для поддержания формы клетки, псевдоподиальных реакций. Роль промежуточных филаментов, по-видимому, также заключается в поддержании структуры клетки. Белки цитоскелета составляют несколько десятков процентов от массы клеточного белка. Центриоли представляют собой цилиндрические белковые структуры, расположенные вблизи ядра клеток животных (у растений центриолей нет). Центриоль представляет собой цилиндр, боковая поверхность которого образована девятью наборами микротрубочек. Количество микротрубочек в наборе может колебаться для разных организмов от 1 до 3. Вокруг центриолей находится так называемый центр организации цитоскелета, район в котором группируются минус концы микротрубочек клетки. Перед делением клетка содержит две центриоли, расположенные под прямым углом друг к другу. В ходе митоза они расходятся к разным концам клетки, формируя полюса веретена деления. После цитокинеза каждая дочерняя клетка получает по одной центриоли, которая удваивается к следующему делению. Удвоение центриолей происходит не делением, а путём синтеза новой структуры, перпендикулярной существующей. Центриоли, по-видимому, гомологичны базальным телам жгутиков и ресничек.
Митохондрии — особые органеллы клетки, основной функцией которых является синтез АТФ — универсального носителя энергии. Дыхание (поглощение кислорода и выделение углекислого газа) происходит также за счёт энзиматических систем митохондрий. Внутренний просвет митохондрий, называемый матриксом отграничен от цитоплазмы двумя мембранами, наружной и внутренней, между которыми располагается межмембранное пространство. Внутренняя мембрана митохондрии образует складки, так называемые кристы. В матриксе содержатся различные ферменты, принимающие участие в дыхании и синтезе АТФ. Центральное значение для синтеза АТФ имеет водородный потенциал внутренней мембраны митохондрии. Митохондрии имеют свой собственный ДНК-геном и прокариотические рибосомы, что безусловно указывает на симбиотическое происхождение этих органелл. В ДНК митохондрий закодированы совсем не все митохондриальные белки, большая часть генов митохондриальных белков находятся в ядерном геноме, а соответствующие им продукты синтезируются в цитоплазме, а затем транспортируются в митохондрии. Геномы митохондрий отличаются по размерам: например геном человеческих митохондрий содержит всего 13 генов. Самое большое число митохондриальных генов (97) из изученных организмов имеет простейшее Reclinomonas americana. Наиболее важным отличием эукариот от прокариот долгое время считалось наличие оформленного ядра и мембранных органоидов. Однако к 1970—1980-м гг. стало ясно, что это лишь следствие более глубинных различий в организации цитоскелета. Некоторое время считалось, что цитоскелет свойственен только эукариотам, но в середине 1990-х гг. белки, гомологичные основным белкам цитоскелета эукариот, были обнаружены и у бактерий. Именно наличие специфическим образом устроенного цитоскелета позволяет эукариотам создать систему подвижных внутренних мембранных органоидов. Кроме того, цитоскелет позволяет осуществлять эндо- и экзоцитоз (как предполагается, именно благодаря эндоцитозу в эукариотных клетках появились внутриклеточные симбионты, в том числе митохондрии и пластиды). Другая важнейшая функция цитоскелета эукариот — обеспечение деления ядра (митоз и мейоз) и тела (цитотомия) эукариотной клетки (деление прокариотических клеткок организовано проще). Различия в строении цитоскелета объясняют и другие отличия про - и эукариот — например, постоянство и простоту форм прокариотических клеток и значительное разнообразие формы и способность к её изменению у эукариотических, а также относительно большие размеры последних. Так, размеры прокариотических клеток составляют в среднем 0,5—5 мкм, размеры эукариотических — в среднем от 10 до 50 мкм. Кроме того, только среди эукариот попадаются поистине гигантские клетки, такие как массивные яйцеклетки акул или страусов (в птичьем яйце весь желток — это одна огромная яйцеклетка), нейроны крупных млекопитающих, отростки которых, укрепленные цитоскелетом, могут достигать десятков сантиметров в длину. Разрушение клеточной структуры (например, при злокачественных опухолях) носит название анаплазии. Деление клетки: Деление эукариотических клеток (рис. 3) Рис 3.
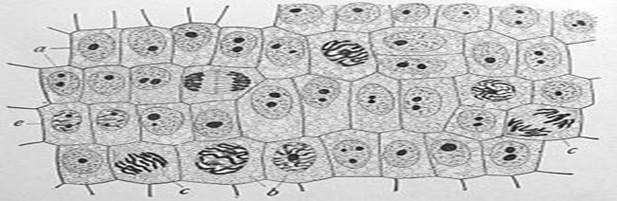
Амитоз — прямое деление клетки, происходит в соматических клетках эукариот реже, чем митоз. В большинстве случаев амитоз наблюдается в клетках со сниженной митотической активностью: это стареющие или патологически измененные клетки, часто обреченные на гибель (клетки зародышевых оболочек млекопитающих, опухолевые клетки и другие). При амитозе морфологически сохраняется интерфазное состояние ядра, хорошо видны ядрышко и ядерная оболочка. Репликация ДНК отсутствует. Спирализация хроматина не происходит, хромосомы не выявляются. Клетка сохраняет свойственную ей функциональную активность, которая почти полностью исчезает при митозе. Таково, например, деление макронуклеусов многих инфузорий, где без образования веретена происходит сегрегация коротких фрагментов хромосом. При амитозе делится только ядро, причём без образования веретена деления, поэтому наследственный материал распределяется случайным образом. Отсутствие цитокинеза приводит к образованию двуядерных клеток, которые в дальнейшем не способны вступать в нормальный митотический цикл. При повторных амитозах могут образовываться многоядерные клетки.
Митоз (от греч. μιτος — нить) — непрямое деление клетки, наиболее распространённый способ репродукции эукариотических клеток, один из фундаментальных процессов онтогенеза. Митотическое деление обеспечивает рост многоклеточных эукариот за счёт увеличения популяции тканевых клеток. Биологическое значение митоза заключается в строго одинаковом распределении хромосом между дочерними ядрами, что обеспечивает образование генетически идентичных дочерних клеток и сохраняет преемственность в ряду клеточных поколений. Дробление оплодотворённого яйца и рост большинства тканей у животных также происходит путём митотических делений. На основании морфологических особенностей митоз условно подразделяется на: профазу, прометафазу, метафазу, анафазу, телофазу. Продолжительность митоза в среднем составляет 1—2 часа. В клетках животных митоз, как правило, длится 30—60 минут, а в растительных — 2—3 часа. Клетки человека за 70 лет суммарно претерпевают порядка 1014 клеточных делений.
Мейоз (от греч. meiosis — уменьшение) или редукционное деление клетки — деление ядра эукариотической клетки с уменьшением числа хромосом в два раза. Происходит в два этапа (редукционный и эквационный этапы мейоза). Мейоз не следует смешивать с гаметогенезом — образованием специализированных половых клеток или гамет из недифференцированных стволовых. Уменьшение числа хромосом в результате мейоза в жизненном цикле ведёт к переходу от диплоидной фазы к гаплоидной. Восстановление плоидности (переход от гаплоидной фазы к диплоидной) происходит в результате полового процесса. В связи с тем, что в профазе первого, редукционного, этапа происходит попарное слияние (конъюгация) гомологичных хромосом, правильное протекание мейоза возможно только в диплоидных клетках или в чётных полиплоидах (тетра-, гексаплоидных и т. п. клетках). Мейоз может происходить и в нечётных полиплоидах (три-, пентаплоидных и т. п. клетках), но в них, из-за невозможности обеспечить попарное слияние хромосом в профазе I, расхождение хромосом происходит с нарушениями, которые ставят под угрозу жизнеспособность клетки или развивающегося из неё многоклеточного гаплоидного организма. Этот же механизм лежит в основе стерильности межвидовых гибридов. Определённые ограничения на конъюгацию хромосом накладывают и хромосомные мутации (масштабные делеции, дупликации, инверсии или транслокации). Прокариотическая клетка. Прокариоты (от лат. pro - перед, до и греч. κάρῠον - ядро, орех) - организмы, не обладающие, в отличие от эукариот, оформленным клеточным ядром и другими внутренними мембранными органоидами (за исключением плоских цистерн у фотосинтезирующих видов, например, у цианобактерий). Единственная крупная кольцевая (у некоторых видов - линейная) двухцепочечная молекула ДНК, в которой содержится основная часть генетического материала клетки (так называемый нуклеоид) не образует комплекса с белками-гистонами (так называемого хроматина). К прокариотам относятся бактерии, в том числе цианобактерии (сине-зелёные водоросли), и археи. Потомками прокариотических клеток являются органеллы эукариотических клеток - митохондрии и пластиды.
Деление прокариотических клеток. Прокариоти́ческие клетки делятся надвое. Сначала клетка удлиняется, в ней образуется поперечная перегородка. На завершающем этапе дочерние клетки расходятся. Отличительной чертой деления прокариотических клеток является непосредственное участие реплицированной ДНК в процессе деления. Обычно прокариотические клетки делятся с образованием двух одинаковых по размеру дочерних клеток, поэтому этот процесс ещё иногда называют бинарным делением. В связи с тем, что в подавляющем большинстве случаев прокариотические клетки имеют клеточную стенку, бинарное деление сопровождается образованием септы — перегородки между дочерними клетками, которая затем расслаивается посередине.
Методы изучения клетки: Световой микроскоп. В изучении клеточной формы и структуры первым инструментом был световой микроскоп. Его разрешающая способность ограничена размерами, сравнимыми с длиной световой волны (0,4–0,7 мкм для видимого света). Однако многие элементы клеточной структуры значительно меньше по размерам. Другая трудность состоит в том, что большинство клеточных компонентов прозрачны и коэффициент преломления у них почти такой же, как у воды. Для улучшения видимости часто используют красители, имеющие разное сродство к различным клеточным компонентам. Окрашивание применяют также для изучения химии клетки. Например, некоторые красители связываются преимущественно с нуклеиновыми кислотами и тем самым выявляют их локализацию в клетке. Небольшая часть красителей – их называют прижизненными – может быть использована для окраски живых клеток, но обычно клетки должны быть предварительно зафиксированы (с помощью веществ, коагулирующих белок) и только после этого могут быть окрашены. Перед проведением исследования клетки или кусочки ткани обычно заливают в парафин или пластик и затем режут на очень тонкие срезы с помощью микротома. Такой метод широко используется в клинических лабораториях для выявления опухолевых клеток. Помимо обычной световой микроскопии разработаны и другие оптические методы изучения клетки: флуоресцентная микроскопия, фазово-контрастная микроскопия, спектроскопия и рентгеноструктурный анализ.
Электронный микроскоп. Электронный микроскоп имеет разрешающую способность ок. 1–2 нм. Этого достаточно для изучения крупных белковых молекул. Обычно необходимо окрашивание и контрастирование объекта солями металлов или металлами. По этой причине, а также потому, что объекты исследуются в вакууме, с помощью электронного микроскопа можно изучать только убитые клетки.
Авторадиография. Если добавить в среду радиоактивный изотоп, поглощаемый клетками в процессе метаболизма, то его внутриклеточную локализацию можно затем выявить с помощью авторадиографии. При использовании этого метода тонкие срезы клеток помещают на пленку. Пленка темнеет под теми местами, где находятся радиоактивные изотопы.
Центрифугирование. Для биохимического изучения клеточных компонентов клетки необходимо разрушить – механически, химически или ультразвуком. Высвобожденные компоненты оказываются в жидкости во взвешенном состоянии и могут быть выделены и очищены с помощью центрифугирования (чаще всего – в градиенте плотности). Обычно такие очищенные компоненты сохраняют высокую биохимическую активность.
Клеточные культуры. Некоторые ткани удается разделить на отдельные клетки так, что клетки при этом остаются живыми и часто способны к размножению. Этот факт окончательно подтверждает представление о клетке как единице живого. Губку, примитивный многоклеточный организм, можно разделить на клетки путем протирания сквозь сито. Через некоторое время эти клетки вновь соединяются и образуют губку. Эмбриональные ткани животных можно заставить диссоциировать с помощью ферментов или другими способами, ослабляющими связи между клетками. Американский эмбриолог Р.Гаррисон (1879–1959) первым показал, что эмбриональные и даже некоторые зрелые клетки могут расти и размножаться вне тела в подходящей среде. Эта техника, называемая культивированием клеток, была доведена до совершенства французским биологом А.Каррелем (1873–1959). Растительные клетки тоже можно выращивать в культуре, однако по сравнению с животными клетками они образуют большие скопления и прочнее прикрепляются друг к другу, поэтому в процессе роста культуры образуются ткани, а не отдельные клетки. В клеточной культуре из отдельной клетки можно вырастить целое взрослое растение, например морковь.
Микрохирургия. С помощью микроманипулятора отдельные части клетки можно удалять, добавлять или каким-то образом видоизменять. Крупную клетку амебы удается разделить на три основных компонента – клеточную мембрану, цитоплазму и ядро, а затем эти компоненты можно вновь собрать и получить живую клетку. Таким путем могут быть получены искусственные клетки, состоящие из компонентов разных видов амеб. Если принять во внимание, что некоторые клеточные компоненты представляется возможным синтезировать искусственно, то опыты по сборке искусственных клеток могут оказаться первым шагом на пути к созданию в лабораторных условиях новых форм жизни. Поскольку каждый организм развивается из одной единственной клетки, метод получения искусственных клеток в принципе позволяет конструировать организмы заданного типа, если при этом использовать компоненты, несколько отличающиеся от тех, которые имеются у ныне существующих клеток. В действительности, однако, полного синтеза всех клеточных компонентов не требуется. Структура большинства, если не всех компонентов клетки, определяется нуклеиновыми кислотами. Таким образом, проблема создания новых организмов сводится к синтезу новых типов нуклеиновых кислот и замене ими природных нуклеиновых кислот в определенных клетках.
Слияние клеток. Другой тип искусственных клеток может быть получен в результате слияния клеток одного или разных видов. Чтобы добиться слияния, клетки подвергают воздействию вирусных ферментов; при этом наружные поверхности двух клеток склеиваются вместе, а мембрана между ними разрушается, и образуется клетка, в которой два набора хромосом заключены в одном ядре. Можно слить клетки разных типов или на разных стадиях деления. Используя этот метод, удалось получить гибридные клетки мыши и цыпленка, человека и мыши, человека и жабы. Такие клетки являются гибридными лишь изначально, а после многочисленных клеточных делений теряют большинство хромосом либо одного, либо другого вида. Конечный продукт становится, например, по существу клеткой мыши, где человеческие гены отсутствуют или имеются лишь в незначительном количестве. Особый интерес представляет слияние нормальных и злокачественных клеток. В некоторых случаях гибриды становятся злокачественными, в других нет, т.е. оба свойства могут проявляться и как доминантные, и как рецессивные. Этот результат не является неожиданным, так как злокачественность может вызываться различными факторами и имеет сложный механизм.

