 2015-05-20
2015-05-20 1825
1825Буынның сілемейлі қабығы қабынуын артрит дейді. Артрит өзбетінше ауру болуы немесе жүйелік аурулардың көріністері ретінде болуы ықтимал.
Пайда болу себептеріне қарай:
√ жұқпалармен байланысты артриттерді, (бактериялардың тікелей сілемейлі қабықты бүліндіруінен, мәселен, жіті іріңді артрит, соз ауруылық артрит, туберкулездік артрит, мерез ауруылық артрит т.б.);
√ иммундық дерттік артриттерді (жіті құздамалық шабуыл кезіндегі артрит, Шенлейн-Генох ауруы кезіндегі немесе екпелерден кейінгі т.б. артриттер);
√ бейинфекциялық ықпалдардан дамитын артриттерді (қан аурулары, подагра кездеріндегі артриттер, паранеоплазиялық артрит);
√ белгісіз себептерден дамитын артриттерді (құздама тәрізді артрит) – ажыратады.
Құздама тәрізді артрит
Құздама тәрізді артрит белгісіз себептердендамитын аутоиммундық дерт. Содан буындардың (негізінен сілемейлі қабығының) және ішкі ағзалардың жүйелік сүлде қабынуы дамиды.
Этиологиясы әлі толық анықталмаған. Бүгінгі таңда жорамал бойынша оның пайда болуында:
√ тұқым қуалаушылыққа бейімділіктің маңызы бар. Жақын туыстардың арасында бұл дерт 16 есе жиі кездеседі. Құздама тәрізді артрит дамуы мен HLA тін үйлесімділік жүйесінің DR1, DR4, DRW4, DW4, DW14 антигендерінің арасында тікелей байланыс болатыны белгілі. Организмнің иммундық жауабына қатысы бар бұл антигендер әртүрлі микробтарға иммунитеттің жасушалық және сұйықтық тетіктерін әсерлендіріп артрит дамуына әкелуі мүмкін;
√ вирустардың (Эпштейн-Барр вирусы, ретровирустар, қызамық, ұшық вирустары; цитомегаловирус т.б.) және микобактериялардың маңызы бар.
Вирус синовиоциттерді тікелей бүліндіруі мүмкін немесе вирус жұқтырған лейкоциттер буын қуысына өтуден олардан босап шыққан вирус синовиоциттермен фагоциттелініп сілемейлі қабықты бүлдіреді – деп есептеледі.
Патогенезі. Құздама тәрізді артрит пайда болуында, Т-супрессор лимфоциттердің қызметі төмендеуімен көрінетін, жүре пайда болған иммундық тапшылықтан дамитын аутоиммундық жайылмалы сүлде қабынудың маңызы бар. Әлі анықталмаған себепкер ықпал макрофагтармен, CD4+Т-лимфоциттерімен әрекеттесіп иммундық серпіліс туындатады. Макрофагтар, тін үйлесімділігінің II-түрдегі үлкен кешенімен бірігіп, антигенді Т-хелпер жасушаларына таныстырады. Макрофагтар өндіретін IL-1, IL-12 CD4+ хелпер жасушаларын әсерлендіріп, олардың өсіп-өнуін арттырады. Содан буынның сілемейлі қабығына Т-хелпер лимфоциттері, плазмалық жасушалар, макрофагтар сіңбеленеді. Әсерленген CD4+ хелпер жасушалары IL-2 өндіріп В-лимфоциттерінің өсіп-өнуін және олардың плазмалық жасушаларға нақтылануын сергітеді. Плазмалық жасушалар сілемейлі қабықта өзгерген және өзара жабысқан IgG өндіре бастайды. Организмнің иммундық жүйесі бұл иммундық глобулинді бөтен текті антиген ретінде танып, сілемейлі қабықтың, лимфалық түйіндердің, көкбауырдың плазмалық жасушаларында оған қарсы IgM түріндегі антиденелер өндіреді. Бұл антиденелер құздамалық фактор делінеді. Ол құздама тәрізді артритпен ауыратын науқастардың 70-80%-да анықталады. Құздамалық фактор шамалы мөлшерде дені сау адамдарда да, жүйелі қызыл жегі, аутоиммундық сүлде гепатит, Шегрен синдромы, гемобластоз, өспе өсуі кездерінде де анықталуы ықтимал. Кейде құздама тәрізді артрит кезінде қанда басқа (ДНҚ молекуласына, жасуша ядросына, қан жасушаларына, коллагеннің II түріне т.б.) аутоантиденелер де табылады.
Өзара жабысқан IgG құздамалық фактормен әрекеттесуінен иммундық кешен құрылады. Бұл иммундық кешенді сілемейлі қабықтың макрофагтары мен нейтрофилдері фагоциттейді. Содан нейтрофилдер бүлініске ұшырап, олардан лизосомалық ферменттер, қабыну медиаторлары (гистамин,, серотонин, кининдер, простагландиндер мен лейкотриендер т.б.) босап шығады. Бұлар сілемейлі қабық пен шеміршекті бүліндіреді. Сонымен қатар, иммундық кешендер, тромбоциттердің агрегациясын, микротромбтар қалыптасуын туындатып, микроциркуляцияны бұзады. Бұл кезде буын антигендеріне қарсы аутоантиденелер ұдайы өндірілуіне байланысты қабыну сүлде түрде өтеді. Дәнекер тіннің және басқа ағзалар мен жүйелердің бүліністері иммундық кешендердің әсерінен васкулит дамуымен байланысты болады – деп есептеледі.
Құздама тәрізді артрит даму патогенезінде цитокиндердің маңызы өте зор. Буын тіндерінде және сұйығында IL-1, IL-6, өспені жоятын альфа-фактор, гранулоциттер мен макрофагтардың колония құруын сергітетін фактор т.б. цитокиндер көптеп анықталады. Бұл цитокиндер сілемейлі қабық жасушаларында, макрофагтар мен фибробластарда өндіріледі және келесі жолдармен қабыну дамуын күшейтеді:
√ қабыну дамытатын простагландер түзілуін арттырады;
√ сілемейлі қабық жасушаларында адгезиялық молекулаларды (селектиндер, интегриндер, тамыр жасушалық адгезиялық молекулаларды -VCAM-1, жасуша аралық адгезиялық 1 және 2 молекулаларды - ICAM-1 және 2) әсерлендіріп бұл қабыққа лимфоциттер, моноциттер мен макрофагтар жабысуын күшейтеді;
√ шеміршектер мен сүйек тінін ыдырататын (коллагеназа, желатиназа т.б.) ферменттердің белсенділігін көтереді;
√ әртүрлі жасушалардың мембранасында тін үйлесімділігі үлкен кешені II түрінің молекулаларын қатты өршітіп, аутоиммундық үрдіс дамуына мүмкіншілік туындатады;
√ гранулоциттер мен макрофагтардың колония құруын сергітетін фактордың әсерінен нейтрофилдер мен макрофагтарда липидтердің асқын тотығуы қатты көтеріледі; иммундық кешендерді фагоциттейтін лейкоциттер буын қуысына шығады;
√ сілемейлі қабықта жаңа тамырлар құрылуын (ангиогенезді) күшейтіп буын қуысына лейкоциттердің шығуын арттырады.
Сонымен бірге, IL-1 және өспені жоятын бета-фактор IL-6 өндірілуін қатты күшейтеді. Ол өз алдына бауыр жасушаларында жіті кезеңдік нәруыздар (С-реактивті нәруыз, фибриноген т.б.) түзілуін көбейтеді, буын сүйектерінде остеопороз дамуын, В-лимфоциттерінің плазмалық жасушаларға нақтылануын және құздамалық фактор түзілуін туындатады.
Осылармен қатар, құздама тәрізді артрит дамуында эндотелий жасушаларының қатысуы да өте маңызды орын алады. Олар цитокиндер (IL-1, 6, 8, тромбоциттік өсу факторы т.с.с.) өндіріп иммундық қабыну дамуын реттейді, қабынудың жіті және сүлде кезеңдеріне қатысады.
Қорыта келгенде, құздама тәрізді артрит кезінде аутоиммундық қабыну дамуынан буынның бүлінуі сілемейлі қабықтың қабынуымен (синовит) басталып, артынан буындарда белсенді өсіп-өнетін фибробластардан, лимфоциттерден, макрофагтардан және жаңадан пайда болған көптеген қан тамырларынан тұратын және қатты өсіп-өнетін қызылшақа (грануляциялық) тін қалыптасады. Осы жағдайды паннус (лат. pannus-жамау, мата кесіндісі) дейді (- сурет). Бұл тін қарқынды өсіп сілемейлі қабықтан шеміршекке тарайды және оны коллагеназа т.б. протеолиздік ферменттерімен ыдыратады. Содан буын ішіндегі шеміршек біртіндеп жоғалады, оның орнында қызылшақа тін болуынан шорбуындық дамиды. Буын қабы, байламдары, сіңірлерінің сүлде қабынуынан буынның сыртқы пішіні өзгереді, шалатаюы, сіресіп қалуы байқалады.

- сурет. Құздама тәрізді артрит кезіндегі жасушалардың қатты өсіп-өнуінен қызылшақа тін (паннус) қалыптасуы.
Құздама тәрізді артрит даму жолдарында аутоиммундық құбылыстар дерттің бастапқы сатыларында маңызды орын алып, соңғы кезеңдерінде иммундық емес тетіктердің (паннустың өсу қабілеті, буын шеміршектеріне жайылуы және оларды бүлдіруі) маңызы артады – деген пікір кездеседі.
Қорыта келе, құздама тәрізді артрит даму патогенезі – сызбанұсқа мен - суретте келтірілген.
Бұл артрит кезінде остеопороз дамиды. Бұл кезде моноциттер мен макрофагтарда өндірілетін IL-1, IL-6, IL-11, өспені жоятын фактор т.б. цитокиндердің сүлде қабыну дамытуы және остеокластарды әсерлендіруі, буын қимылының азаюымен қабаттасып остеопороз дамытады.
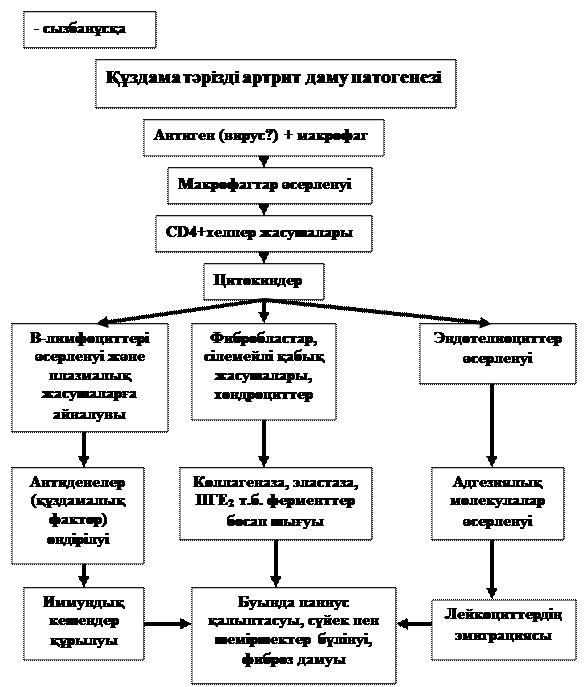
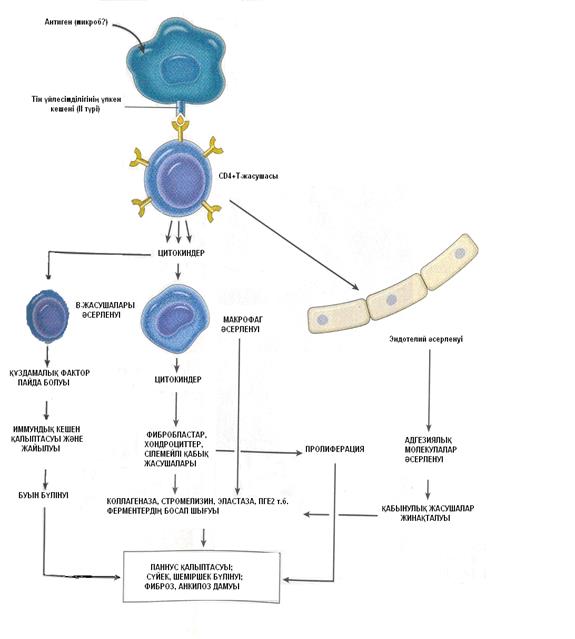
- сурет. Құздама тәрізді артриттің патогенезі
Бұлшықеттердің біртектес бүліністері
Бұлшықеттердің бүліністері жүре пайда болған және туа біткен болуы мүмкін.
Бұлшықеттердің жүре пайда болған бүліністері
Адамның қартаю кездерінде шапшаң ет талшықтарының жүйкелену белсенділігі төмендеп, ол шабан ет талшықтарында көтеріледі. Бұл кездерде шапшаң және шабан ет талшықтарының арасында өздеріне тән зат алмасулық және функциялық ерекшеліктер біртіндеп жоғалады. Дегенмен шапшаң ет талшықтары шабан ет талшықтарына қарағанда ертерек қартаяды.
Қарттардың бұлшықеттерінде оттегіні пайдалану екі есеге жуық, АТФ мөлшері 25-35%-ға, креатинфосфаттың деңгейі 40-50%-ға азаяды. Сонымен қатар, оларда бос креатин мен АДФ мөлшері көбейеді. Бұндай бұлшықет жасушаларының митохондрийларында сукцинатдегидрогеназа ферментінің белсенділігі төмен болады.
Қартайған бұлшықет талшықтарында метаболизм мен функциялық бүзылыстардың пайда болу негізінде синапс алды құрылымдарынан ацетилхолин босап шығуы азаюы, синапстық саңылауларда оның ыдыратылуы бұзылуы, түйіспе соңы мембранада холинорецепторлардың тығыздығы мен иондарға өткізгіштігі көтерілуі маңызды орын алады. Бұндай жағдайларда күрт деполяризацияның деңгейі көтеріледі, әрекеттік потенциалдың ауытқу аумағы азаяды және оның ұзақтығы ұзарады. Сондықтан бұлшықет талшықтары шапшаң жиырылу қабілетінен айырылады да негізінен шабан қызмет атқарады. Қаңқа бұлшықеттерінің қартаюға байланысты өзгерістері қарттарда жиі орын алатын гипокинезияның әсерінен ушығып кетуі ықтимал. Осыдан бұлшықеттердің жиырылу күші төмендеуі, ет талшықтарының семуі және олар мен жүйкелер қосылыстарының бүліністері байқалады.
Гиподинамия ұзақ төсек тартып жатқан науқастарда, ғарышта ұшып жүрген ғарышкерлерде, ұдайы отырып жұмыс істейтін адамдарда бұлшықеттердің межеқуаты бір деңгейде сақталуымен байқалады. Бұндай жағдайларда бұлшықеттерден, тарамыстар мен буындардан мыйға бағытталалатын серпіндер қатты азаяды да, мыйдың торлы құрылымынан жұлынның нейрондарын сергітетін ықпалдар әлсірейді. Осыдан шапшаң ет талшықтарына қимылдық нейрондардың әсері азаяды, бұлшықеттердің межеқуаты төмендейді, оларға жүйкелердің нәрленістік әсері бұзылады. Бұлшықеттерде энергия шығындалуы азаяды, артериолалар мен көктамырлар арасындағы ашық анастомоздар көбейеді, қылтамырларда қан ағуы баяулайды. Сол себепті ет талшықтарында гипоксия дамып, тотығулық фосфорлану бұзылудан кесек қуатты энергия түзілуі кемиді. Энергия тапшылығынан кальций насостарының қызметі әлсірейді, жасушаларда ДНҚ, РНҚ және нәруыздар түзілуі азаяды, анаболизмдік үрдістерден катаболизмдік үрдістер басым бола бастап, ет талшықтары (әсіресе шапшаң ет талшықтары) нәруыздарының ыдырауы күшейеді. Ол өз алдына миофибрилдердің, шапшаң ет талшықтарында саркомерлердің азаюына және олардың атрофиясына, бұлшықеттердің қозымдығылы мен жиырылғыштық қабілеттері төмендеуіне әкеледі.
Қимылдық мүшелерді гипспен таңып тастағанда, буындар сіресіп қатып қалғанда т.б. жағдайларда бұлшықеттердің қимылы ұзақ болмайды. Таңып бекіту екі түрлі жағдайда болуы мүмкін. Біріншісі, бекітілген мүшенің бұлшықеттері қысқартылған жағдайда, екіншісі ұзартылған жағдайда.
Аяқ-колды бұлшықеттері қысқартылған жағдайда таңып тастаудан физиологиялық тыныштық кезінде олардың рецепторлық құралдарының белсенділігі төмендейді, бұлшықеттердің ерікті қозғалыстарында олар бір ұзындықта ғана жиырылады. Осындай жағдайларда бекітілген бұлшықеттерді жүйкелейтін жұлынның мотонейрондарының белсенділігі қатты әлсірейді, қимылдық жүйке талшықтары жұқарады, жүйке аяқшалары жыртылып бүлінеді, постсинапстық қатпарлар ашылып қалады. Бұл өзгерістер жүйкелік трофикалық әсерлердің бұзылыстарымен, ет талшықтарының соңғы табақшалары аумағында жүйке аяқшаларының өсіп кетуімен қабаттасады. Таңып бекітудің бірінші аптасында бұлшықет тінінде тотығулық үрдістердің қарқыны төмендейді, натрий мен хлор иондарының мөлшері көбейеді, РНҚ, нәруыздар түзілу деңгейі азаяды және бұлшықет жасушаларының қайта нақтылануы басталады. Бұл кезде гликолиздік шапшаң ет талшықтарында аэробтық тотығу күшейіп, анаэробтық гликолиз төмендейді. 2-3 аптаның ішінде нәруыздар түзілуі азаяды, бар саркомерлердің 40% жуығы жоғалады, ет талшықтарының көлемі кішірейеді, негізінен шапшаң бұлшықеттің семуі байқалады. Осылармен қатар миозиннің АТФазалық белсенділігі және миофибрилдердің Са2+-тәуелді АТФазасының белсенділігі төмендейді, саркоплазмалық торшалардың Са2+ иондарын сорып алу қызметі көтеріледі.
Таңып бекітудің 1-аптасында шабан ет талшықтары айтарлықтай өзгерістерге ұшырамайды. Ал, 5-6 аптадан кейін оларда тіндік тыныс төмендеп, анаэробтық гликолиз күшейеді, сүт қышқылы түзілуі артады, АТФ мөлшері азаяды. Сонымен қатар нәруыздардың түзілуі кеміп, олардың ыдырауы артады, ет талшықтарының атрофиясы дамиды, саркомерлер азаяды, лизосомалық протеазалардың белсенділігі көтеріледі. Осыдан шабан бұлшықеттердің көлемі кішірейеді. Ет талшықтарында миозиннің АТФазалық белсенділігі және миофибрилдердің Са2+-тәуелді АТФазасының белсенділігі төмендейді.
Қимылдық дене мүшелерін бұлшықеттері ұзартылған жағдайда таңып бекіту кездерінде афференттік серпіндердің тарауы, қимылдық нейрондардың электр өндіру қасиеті, ет талшықтарына жүйкенің нәрленістік қызметі барлығы сақталып қалады. Бұндай жағдайда таңып бекітудің 2-3 аптасында 20%-ға дейін жаңа саркомерлер құрылып, ет талшықтарының керіліп созылу дәрежесі кішірейеді. Бұл кезде шапшаң ет талшықтарының атрофиясы өте баяу дамиды және бірнеше айлар бойы олардың атқаратын міндеттемелері қалыпты деңгейде сақталады. Шабан ет талшықтары бекітілген бұлшықеттің барлық бойында гипертрофияға ұшырайды.
Өндірістік жағдайларда, спортпен шұғылданғанда т.с.с әрекеттерде ұзақ ауыр қол жұмыстарын атқарудан гипердинамия байқалады. Ол жұмыс атқаратын бұлшықеттердің жиырылғыштық және метаболизмдік қасиеттерінің сатылы түрде өзгерістерімен сипатталады.
Бірінші сатысы жұмыс атқаратын бұлшықеттердің жиырылғыштық қабілеті көтерілу сатысы делінеді. Бұл кезде белсенді қимылдық бірліктердің саны көбейеді, олардың әрекеттері бір мезгілдендіріледі, электромиограммада серпіндердің жиілігі мен ауытқу аумағы ұлғаяды, бұлшықеттердің жиырылу күші ең ұлкен деңгейге жетеді. Бұлшықеттердің жиырылғыштық қабілетінің көтерілуі оттегіні, гормондарды, жүйкелік медиаторларды және тотығатын заттарды артық пайдалану арқылы қамтамасыз етіледі. Бұл сатыда катехоламиндер ет талшықтарында Na+, K+ иондарының белсенді тасымалдануын күшейтеді, мембраналық әлеуетті (потенциалды) мөлшермен 10%-ға арттырады. Осымен бір мезгілде қаннан глюкозаның сорылуы мен жасуша ішілік гликоген қорының, әсіресе шапшаң ет талшықтарында ыдырауы артады. Глюкоза тотығуы кезінде пирожүзім қышқылы карбоксил тобынан ажырап, Кребс оралымында пайдаланылатын ацетил-КоА-ға айналады. Бұл саты бұлшықет талшықтарында гликоген қоры таусылғанша болады. Бұлшықеттердің жиырылғыштық қабілетінің көтерілуіне жағымсыз жағдай болып бұлшықеттің ишемиясы есептеледі. Бұндай жағдайда жұмыс атқаратын бұлшықеттерде АТФ және креатинфосфат түзілуі үшін заттардың алмасуы ауыр гликолиздік жолмен жүруінен сүт қышқылы көптеп түзіледі. Оның мөлшері 100-150 ммоль/л-ге жеткенде бұлшықеттердің жиырылғыштығы кеміп кетеді. Өйткені ишемия кезінде бұлшықеттердің жиырылғыштық қабілетін кемітетін ықпал болып, тотығатын заттың таусылуы емес, Н+ иондарының, АДФ мен бейорганикалық фосфордың деңгейі көбеюі есептеледі.
Бұлшықеттердің жиырылу қабілетінің бәсеңсу сатысы оларда гликоген қорының таусылуынан, қанда глюкоза азаюынан және оның ет талшықтарына жеткіліксіз түсуінен дамиды. Бұлшықет, әсәресе шапшаң ет, талшықтарында лактаттың мөлшері көбейіп, метаболизмдік ацидоз дамиды. Осыдан фосфофруктокиназа, креатинкиназа ферменттерінің белсенділігі төмендеп, кесек қуатты энергиялық қоспалардың байқалады. Креатинфосфат пен АТФ мөлшері екі еседен астам төмендейді, АДФ фосфорлану жылдамдығы баяулайды. Бұлшықет талшықтарында энергияның тапшылығы майлардың бета-тотығу жолы есебінен жартылай толықтырылады. Осымен бір мезгілде жұмыс атқарып тұрған бұлшықеттерде аланинаминотрансферазаның қатысуымен аланин түзілуі күшейеді, аминқышқылдарының дезаминденуі және кетоқышқылдардың тотығуы көтеріледі. Қан сұйығында нәруыздық емес азоттың, зәрнәсіл мен зәр қышқылының деңгейі көбейеді. Энергия тапшылығына осылай икемделудің нәтижесінде бұлшықет жасушаларында ацетил-КоА көптеп жинақталады, сонымен қатар эндогендік фосфолипазаның белсенділігі көтерілуінен жасуша мембраналарының құрылымы бүлінеді.Ацетил-КоА деңгейі көбеюі пируватдегидрогеназа ферментінің белсенділігін бәсеңсітеді де карбоксилазаны әсерлендіреді. Осыдан көмірсуларының тотығуы тежеледі, қан сұйығында лактаттың деңгейі азаяды, кетондық денелер көбейеді. Зат алмасуларының бұзылыстарына катехоламиндер мен глюкокортикоидтар түзілуінің тежелуі сәйкес келеді. Ол өз алдына тотығулық үрдістердің қарқынын және ет талшықтарының мембраналарында орналасқан иондық насостардың белсенділігін одан сайын төмендетеді. Саркоплазманың қышқылдануы мен иондардың тасымалдануы бұзылыстары жинақталып тропониннің Са2+-байланыстыратын орталықтарының иондарға үйірлігін төмендетеді, саркоплазмалық торшаларда Са2+ иондары көбейіп, олардың босап шығуы тежеледі, Т-жүйесінде әрекеттік потенциал өткізілуі бұзылады. Осымен бір мезгілде жүйкелік медиатордың шығарылуы үдемелі азаюдан ет талшықтарының тітіркену шегі көтеріледі, соңғы табақшаларда мембраналық потенциалдың жиілігі сирейді. Бұл кезде синапс соңылық мембрананың соңғы табақша потенциалын туындату қабілеті төмендейді, бірақ олардың ацетилхолинге сезімталдығы толығынан сақталып қалады. Өйкені жұлынның қимылдық жүйкелік орталықтарының қозымдылығы төмендеген. Бұл кезде бұлшықеттердің жұмыс атқаруға энергия шығындауы қатты көтеріледі, қимылдық қозғалыстарға рефлекстік түрде қосымша бұлшықеттер жұмылдырылады. Осыдан қимылдық қозғалыстардың атқарылуы кездерінде жекелеген ет талшықтары өзара үйлеспей жиырылады. Бұлшықеттердің жиырылу күші бастапқы деңгейінен айтарлықтай төмендейді. Бұлшықеттердің жиырылу қабілетінің бәсеңсу сатысы қанайналым және тыныс алу жүйелерінің белсенділігі артуымен қабаттасады.
Бұлшықеттердің қатты қажуы тым ауыр физикалық жүктемелердің (әбден шаршағанша жүріру, ұзақ тырыспа селкілдек ұстамасы ұстауы т.с.с.) нәтижесінде байқалады. Ол бұлшықеттерге ұзақ жеңіл жүктемелерге қарағанда қысқа қарқынды жүктемелер кездерінде тезірек дамиды. Жедел қатты қажу әдетте спортшылар тым ауыр жүктемелер орындағаннан кейін ілезде көрінеді. Оның ұзақтығы 2 сағатқа дейін созылады, бұл кезде бұлшықеттер сіресіп, олардың жиырылу күші азаяды және оларда ауыру сезімі пайда болады. Егер тым ауыр жүктеме қатты қажу басталғанша болса, онда бұлшықеттердің функциялық жағдайы тез арада қалпына келеді. Қатты қажу дамуына дейін созылған тым ауыр жүктеменің нәтижесінде митохондрийлардың (әсіресе шапшаң ет талшықтарында) үдемелі бүліністері, миофиламенттерде (миолиз және олардың өзара ажырап кетуі сияқты) бүліністік өзгерістері, гликоген мен креатинфосфаттың қайта түзілуі қатты баялауы, саркоплазмада детрит пайда болуы және оның аластануы бұзылуы, миобластардың пайда болуы байқалады. Ет жасушаларының айналасындағы көрші жасушалардың митоздық бөлінуі артып кетеді. Бұл кезде бүлінген бұлшықеттердің қалпына келуі отыз тәуліктен астам уақытты қажет етеді.
Бұлшықеттердің сүлде ишемиясы жүрек қызметінің сүлде жеткіліксіздігі, гиподинамия, гиповолемия кездерінде, әсіресе олардың біріккен әсерлерінен байқалады. Бұлшықет тінінде қан ағымының және зат алмасулары ыдырау өнімдерінің одан аластануы шектелуі нәтижесінде ет талшықтары жиырылуына қажетті энергия түзілуі азаяды. Бұл кезде ет талшықтарының митохондрийларында креатинфосфокиназа ферментінің қатысуымен жүретін креатиннің фосфорлануы әлсірейді және креатинфосфаттың миофибрилдерге тасымалдануы төмендейді. Бұлшықет жиырылуы энергиямен толық қамтамасыз етілмеуінен ет тінінде гликогенолиз күшейіп, гликогеннің қоры азаяды. Бұлшықеттерде нәруыздардың ыдырауы артады, сүт қышқылы жиналады, ацидоз дамиды, фосфофруктокиназа мен креатинкиназа ферменттерінің белсенділігі төмендейді. Осыдан креатинфосфаттың жеткіліксіздігі дамиды, АДФ фосфорлануы азаяды және сарколемманың өткізгіштігі көтеріледі де жасуша ішінде кальций иондарының мөлшері көбейеді. Ол кальций тәуелді протеазалардың белсенділігін арттырып, миофибрилдердің ыдырауына әкеледі. Ет талшықтарында дистрофия дамуы олардың тіршілігін жойып, дәнекер тіндер өсуін туындатады.
Бұлшықеттердің жіті ишемиясы артериалық тамырлардың эмболиясы немесе қанауды тоқтату үшін бұрау салу кездерінде байқлады. Бұл жағдайларда бұлшықеттердің бүліну дәрежесі олардағы қанайналым тоқтау ұзақтығынан байланысты. Ол екі сағатқа дейін тоқтағанда бұлшықет тінінде оттегінің үлестік қысымы қатты төмендейді, бос май қышқылдарының ыдыратылуы екі еседен астам әлсірейді, анаэробтық гликолиз артып, жасуша ішілік және сыртылық ацидоз дамиды. Бұлшықеттерде қанайналым тоқтағаннан кейін 1-1,5 сағаттан соң сүт қышқылының деңгейі 3-еседен астам көтеріледі. Ет нәруыздарының ыдырауы артып, аденозин өндірілуі мен оның жасушалар сыртына шығарылуы көбейеді. Ишемияға ұшыраған бұлшықеттерде кесек қуатты АТФ, креатинфосфат түзілуі азаяды. Сонымен бірге бұлшықеттерде катехоламиндердің деңгейі төмендейді, оларға жүйкелердің нәрленістік қызметі әлсірейді. Зат алмасуларының бұзылыстары нәтижесінде антиоксиданттық ферменттер түзілуі азаяды, жасушалар мембранасында липидтердің асқын тотықтары жинақталады. Содан кальций-тәуелді АТФаза т.б. тасымалдық ферменттердің белсенділігі төмендейді, мембраналардың, әсіресе сарколемманың, қайтымсыз бүліністері пайда болып, иондардың мембрана арқылы тасымалдануы бұзылады. Калий иондары жасушалардың сыртына шығарылып, олардың ішінде натрий мен хлор иондары жиналады, осмостық қысым көтеріліп, жасушалар ісінеді. Жасушалардың көлемі ұлғаюдан газдардың диффузиясына арақашықтық алшақтайды, жасушаға оттегі, заталмасу өнімдері, ферменттер жеткізілуі азаяды және бұлшықеттердің жиырылу қабілеті жоғалады. Бұл кезде, осындай заталмасулық және функциялық бұзылыстарға қарамай, бұлшықеттердің ультрақұрылымындағы өзгерістер шамалы ғана болады. Бұлшықетте қанайналым қайта қалпына келсе жасушаларда АТФ, креатинфосфат, гликоген және басқа энергия көздері толығынан қалпына келеді. Ишемияның қайтымды сатысында бүлінген бұлшықетте әрекеттік потенциалдың тарауы және ұзақтығы баяулайды, элктромиографияда (ЭМГ) сирек жиілікті толқындар басым болады, шеткері бағытталған эфференттік серпіндердің өткізілуі толығынан сақталып, мыйға бағытталған афференттік серпіндердің өткізілуі сұрыпталып бөгеледі. Осыдан бір бағытта (синергистік) және қарама-қарсы (антагонистік) әсерленетін бұлшықеттерді жүйкелендіретін жұлын орталықтары әрекеттерінің үйлесімі бұзылады.
7 сағаттан астам қанайналым болмауынан ет тінінде зат алмасуларының ауыр бұзылыстары, асқын тотығу үрдістерінің қатты артуы, эндогендік фосфолипаза ферментерінің әсерленуі, жасуша ішілік құрылымдардың бүлінуі және нәруыздардың ыдырауы байқалады. Қайтымсыз бүлінген ет талшықтарында қанайналым қайта дұрысталған жағдайда анаэробтық гликолиз одан сайын артады, ет талшықтарының тіршілігін жоюылуына келетін липидтердің асқын тотығуы қатты үдеп кетеді.
Бұлшықеттердің тікелей бүліністері механикалық (жарақат, қысылу, созылу т.с.с.) ықпалдардан, химиялық бүліндіргіш заттардың (өт қышқылдары, майда еритін заттар – эфир, хлороформ, фторотан т.б.) әсерлерінен байқалады. Бұл бүліністер әсер еткен ықпалдың қарқынына, әсер ету ұзақтығына тікелей байланысты болады және ет талшықтарының әртүрлі құрамбөлшектеріне тарайды. Сарколемманың бүлінісі мембран арқылы Na+, K+ және Cl-иондарының белсенді және енжар тасымалдануы бұзылады, әректтік потенциал қалыптасуы мен оның Т-жүйесіне берілуі әлсірейді. Т-жүйесі қозбауынан саркоплазмалық торшалардың мембранасында электрлік поляризация пайда болмайды, олардан Са2+ иондары саркоплазмаға шығарылмайды. Содан саркоплазмада Са2+ аз болудан ет талшықтарының жиырылу қабілеті төмендейді. Митохондрийлардың бүліністері ерте дамиды. Олардың нәтижесінде макроэргиялық фосфор қосындыларының түзілуі азайып, энергияның тапшылығынан мембраналар арқылы иондардың белсенді тасымалдануы, бұлшықет тінінің жиырылу-босаңсу қызметтері бұзылады. Саркоплазмалық торшалардың бүлінісі липидтердің асқын тотықтары жиналып қалуымен, мембранада асқын тотықтық тесіктердің пайда болуымен, мембрананың липидтік қабаты мен Са2+- тәуелді АТФаза түйісетін аумақта нәруыздар мен липидтердің өзара әрекеттесуінің бұзылыстарымен және Са2+-ді байланыстыратын бөлікшелердің өзгерістерімен сипатталады. Мембрананың Са2+-иондарына енжар өткізгіштігі қатты көтеріледі және Са2+-тәуелді АТФазаның белсенділігі бәсеңсиді, саркоплазмада Са2+-иондарының мөлшері көбейеді. Осыдан ет талшықтары сіресіп жиырылып қалуы, Са2+-тәуелді протеазалардың белсенділігі көтерілуі, миолиз дамуы байқалады. Бұлшықеттің механикалық ықпалдардан жыртылып кетуінен кейін артына ет талшықтарының қалпына келу кезеңі болады. Тіректік мембрананың астында орналасқан ет талшықтарында миобластардың өндірілуі артып, олардың өсіп-өнуінен бұлшықеттің регенерациясы болады.
Бұлшықеттердің иммундық бүліністері. Әртүрлі себептерден (некроз, миолиз) сарколемманың тосқауылдық қызметі бұзылып, оның өткізгіштігі артуынан миоциттердің нәруыздары (тропонин, тропомиозин т.б.) қанға түсуі аутоиммундық үрдістер дамуына әкеледі. Осыдан бұлшықеттерге қарсы аутоантиденелер пайда болуынан бұлшықеттерді бүлдіретін цитотоксиндік аллергиялық серпілістер дамиды.
Бұлшықеттердің метаболизмдік бүліністері толық және жартылай ашығу кездерінде кесек қуатты фосфор қоспаларының түзілуі тапшылығынан жиі дамиды. Бұл кездерде негізінен гликолиздік-тотығулық талшықтар бүлінеді. Гликолиздік талшықтардың митохондрийларында май қышқылдары мен кетондық денелердің тотығу жолдары сақталғандықтан бұлар ашығуға біршама тұрақты болады.
Бұлшықет талшықтарының жергілікті бүліністерімен Ценкер некрозы байланысты дамиды. Ет талшықтарының бүлінісіне жауап ретінде қатты жиырылу аумақтары пайда болады, протофибрилдер ыдырайды, саркоплазмалық торшалар мен Т-жүйесінің бөлікшелері бүлінеді. Осылардың нәтижесінде ет талшықтарында көпіршіктер пайда болады. Ценкер некрозы таралу аумағының көлеміне тікелей байланысты бұлшықеттің жиырылу қабілеті бұзылады.
Организмде глюкокортикоидтардың тым артық өндірілуі немесе бұлшықет тінінде зат алмасуларына қажетті витаминдердің тапшылығы да, жоғарыда келтірілгендермен қатар, бұлшықеттер жүйесі бұзылыстарына әкеледі.
Стероидтық миопатия глюкортикоидтарды көп мөлшерде егу арқылы ұзақ қабылдаудан немесе Иценко-Кушинг ауруы кездерінде организмде осы гормондардың деңгейі ұзақ көтерілуінен дамиды. Ет талшықтарына көп мөлшерде глюкокортикоидтар тым артық түсуінен олар арнайы рецепторлармен көптеп байланысады, ет талшықтары нәруыздары ыдырауын арттырып, олардың түзілуін кемітеді, миоциттердің ішіне Na+ иондары енуін күшейтіп, Na+/К+ арақатынасын көбейтеді. Содан шапшаң гликолиздік талшықтардың жартылай деполяризациясы дамиды, әрекеттік потенциалдың пайда болу межесі және шапшаң бұлшықет талшықтарының жиырылу күші төмендейді.
Е авитамнозы кезінде миопатия дамиды. Оның даму негізінде ет талшықтарында Са2+-өзекшелері мен, Са2+-насостары әрекеттерінің және кальций алмасуының бұзылыстары маңызды орын алады. Бұл кезде саркоплазмалық торшалардың мембранасында липидтердің асқын тотықтары жиналады, мембрананың липидтерімен Са2+-АТФ-азаның түйісу аумағында нәруыздар мен липидтердің өзара әрекеттесуі бұзылады, Са2+-АТФ-азаның белсенділігі төмендейді. Содан саркоплазмалық торша мембраналары бөлшектерінің Са2+-иондарына өткізгіштігі қатты көтеріледі және саркоплазмада иондалған Са2+ мөлшері көбейеді. Осыдан Са2+-тәуелді протеазалардың белсенділігі артады, миофибрилдердің жирылу қабілеті бұзылады, миолиз дами бастайды, тропонинің мөлшері көбейеді. Ет талшықтарында митохондрийлар ісінеді, олардың тарақшалары бөлшектеледі, АДФ молекуласының фосфорлануы азаяды, электрондарды тасымалдайтын жүйенің қызметі бәсеңсиді. Осылардың нәтижесінде бұлшықеттерде дистрофия дамиды.
Бұлшықеттердің туа біткен бүліністері
Бұлшықеттердің жиырылу босаңсу тетіктеріне қажетті энергия қуаты алмасуының туа біткен бұзылыстары болуы ықтимал. Бұндай аурулар бұлшықеттердің әлсіздігімен, дерттік түрде тез қажуымен т.с.с. көрінетін олардың жиырылғыштық қабілеті бұзылыстарымен сипатталады.
Креатинфосфокиназаның туа біткен ақауы анасының Х хромосомасындағы геннің қызметі өзгеруінен және ол ақаудың ішіндегі ұрыққа берілуінен дамиды. Бұл дерттің даму негізінде май қышқылдарының метаболизміне қажетті энергия тасымалдау қызметінің бұзылуы жатады және ол бұлшықеттерде әр түрлі дистрофиялар дамуымен көрінеді. Митохрийлар мен саркоплазмада болатын креатинфосфокиназаның ақауы нәтижесінде оның қатысуымен өтетін креатиннің фосфорлануы болмайды. Содан креатинфосфаттың ет талшықтары арасындағы кеңістіктерге тасымалдануы, одан АДФ молекуласына фосфор қышқылы ауысуы бұзылады, креатиннің ет талшықтары арасындағы кеңістіктерден митохондрийларғы тасымалдануы ауырлайды. Ет талшықтарында креатинфосфат пен АТФ молекуласының тапшылығы пайда болады, қан сұйығынан креатин бұлшықеттерде пайдаланылмай, несеппен көптеп шығарылады.
Мембрана фосфолипидтері құрамының туа біткен бұзылыстары (туа бітен ет дистрофиясы) сарколеммада фосфатидилхолин азаюымен, керісінше, фосфатидилэтаноламин, сфингомиелин, инозитол- және серинфосфатидтер көбеюімен сипатталады. Осылармен бірге ацетилхолинэстераза белсенділігі 2-3 есе көтеріледі. баяу ет талшықтарының сарколеммасында көптеген тесіктер пайда болады. Сондықтан баяу бұлшықеттерде шапшаң талшықтарға қарағанда ауыр дерттік өзгерістер дамиды. Бүлінген бұлшықеттердің сарколеммасының сыртқы беттерінде қан қылтамырлары көбейеді. Сонымен қатар бүлінген ет талшықтары қалыпты жағдайда болмайтын эндоцитоздық қасиет қабылдайды. Ол лизосомалық жүйемен тығыз байланысты болады. Бұндай ет талшықтарында жекелеген жиырылу даму уақыты, оның жартылай босаңсу және толық жиырылу аралық кезеңі ұзарады. Жиырылу қызметінің бұзылуы ет талшықтарында әрекеттік потенциал қалыптасуы бұзылуымен және оның ауытқу мөлшері (амплитудасы) төмендеуімен қабаттасады. Бүлінген бұлшықеттерден мыйға бағытталатын серпіндер сақталып қалады және ет талшықтарының денервацияға жауабы қалыпты бұлшықеттердегідей болады. Сонымен туа біткен бұлшықет дистрофиясы кездерінде афференттік және эфференттік жүйелердің елеулі бұзылыстары байқалмайды.
Дюшеннің жалған гипертрофиялық үдемелі миопатиясы анасының Х хромосомасымен тек ұлдарында дерттік ген бәсеңкі жолмен берілуінен дамитын біріншілік миопатия. Науқас балалардың ет талшықтарында дистрофин нәруызының түзілуі болмайды. Содан сарколеммада құрылымдық, функциялық және ферменттік ауытқулар пайда болады. Бүлінген ет талшықтарында анаэробтық гликолиз ферменттерінің белсенділігі төмендейді және бұлшықет нәруыздарына аминқышқылдарының (тирозин т.б.) енуі артуы изоцитратдегидрогеназа ферментінің белсенділігі көтерілуімен қабаттасады.
Дерттің бастапқы сатысында сарколемманың шағын бөліктерінің жартылай бүліністері байқалады. Артынан бұлшықеттердің шамалы әлсіздігі дамуымен және ет талшықтарының құрылымы мен функциясы біртіндеп қалпына оралады. Кейінгі сатысында ет талшықтарында сарколемманың ауыр бүліністерімен, Zдискілерінің еруімен және митохондрийлардың ыдырауымен қабаттасатын бөлікшелік некроздар пайда болады. Бүлінген мембраналардың өткізгіштігі қатты көтерілуден миоциттердің ферменттері тінаралық сұйыққа, артынан қанға шығарылады. Фагоциттелген тіршілігін жоғалтқан талшықтардың қапшықтарында коллаген жиналады, миобласт тәрізді жасушалар өсіп өнеді. Осыдан бұлшықеттердің зақымданған бөліктерінде жалған гипертрофия дамиды.
Миотония адамның тегіне байланысты сарколемманың функциялық және құрылымдық бүліністерімен сипатталатын бұлшықеттердің тұқымға тарайтын ауруы. Бұл кезде бұлшықеттердің қозымдылығы азаяды, ерікті рефлекстік жиырылудан кейін еттің босаңсуы баяулайды. Бүлінген ет талшықтары мембранасының хлорға өткізгіштігі азаяды, натрий мен калийге өткізгіштігі ұлғаяды. Ет талшықтарының жиырылуы қысқа мерзімді, жоғары жиіліктегі электр серпіндері топтамасының әсерінен болады. Ет талшықтарының жиырылулары амплитудасы мен жиілігі төмендеумен көрінетін оң үшкір толқындардан немесе үш фазалы толқындардан тұрады. Миотония кезінде қимылдық жүйке талшықтары арқылы электр серпіндерінің өткізілу жылдамдылығы азаяды, бүлінген бұлшықеттерде қимылдық бірліктердің саны азаяды. Бірақ сарколеммада холинорецепторлардың орналасу тығыздығы қалыпты деңгейде сақталады.
Бұлшықеттердің жүйкелік бүліністері қимылдық жүйке талшықтары қысылып қалғанында, кесілгенінде немесе жарақаттан үзілгенінде, мый мен жұлын орталықтарының ишемиядан, нейротроптық улармен немесе вирустармен зақымдануынан мотонейрондардың тіршілігін жоғалтқанында байқалады. Қимылдық нейрондардың немесе олардың аксондарының кезкелген бүліністерінде аксоплазмалық тасымалдану тоқталады, жүйкелік-еттік синапстар арқылы нейромедиаторлар мен трофогендер өтпеуінен, бұлшықеттерге жүйкелердің функциялық және нәрленістік әсері болмайды. Осындай жағдайды жүйкесізденулік немесе денервациялық синдром дейді. Бұл кезде шапшаң және баяу бұлшықеттердің жиырылуы әлсірейді және баяулайды, олардың арасында қалыпты жағдайда болатын ферменттік айырмашылықтар жоғалады. Жүйкесізденген бұлшықеттерде ет талшықтарының көлемі, дара жиырылулардың амплитудасы кішірейеді, әрекеттік потенциалдың деңгейі және оның сарколемма арқылы тарауы азаяды. Артынан жүйке талшықтарының ыдырауы, ақырғы табақшалардың бөлшектелуі болады. Осыдан бастап бұлшықеттің жүйкесіздену тетіктері қосылады.
Шапшаң гликолиздік ет талшықтарында аэробтық тотығуға қатысатын ферменттердің түзілуі тежелуден шығып кетеді. Содан анаэробтық гликолиздің бәсеңсуімен бір мезгілде тіндік тыныс және оттегіге мұқтаждық артып кетеді. Бұл кезеңде фосфорлық қосындылардың жаңаруы тездейді, креатинфосфаттың ыдырауы азаяды, холинацилтрансферазаның белсенділігі төмендейдіацетилхолин түзілуі азаяды, жалған холинэстеразаның белсенділігі артады. Тыныштық жағдайда ет талшықтарының калий иондарына өткізгіштігі көтеріледі, қалыпты натрий өзешелерінің саны азаяды, шабан натрий өзекшелері пайда болады. Na+-K+-АТФазаның сырттан енгізілген ацетилхолинге сезімталдығы қатты көтеріледі, өз бетінше жыпылықтайтын әрекеттік потенциал пайда болады. Жүйкесіздену кезінде ферменттер түзілуі артуымен қатар саркоплазмалық торшаларда полипептидтк және олигопептидтік тізбектердің түзілуі көбейеді, Гөлдж аппаратында холинорецепторлардың түзілуі және жинақталуы күшейеді. Бұл рецепторлар артына саркплазмаға тасымалданып, сарколемманың құрамына енеді. Холинрецепторлар синапстардан тыс көптеп орналасуынан ет талшықтарының сезімталдығы тым көтеріліп кетеді. Ет талшықтары, саркоплазмада ацетилхолиннің мөлшері қатты азаюына қарамай, организмнің сұйықтарында айналып жүрген ацетилхолиннің аз мөлшерлеріне жауап қайтара бастайды. Бұндай әдеттен тыс сезімталдық холинорецепторлардың лизосомаларда ыдыратылуы бұзылуына байланысты болуы ықтимал. Ет талшықтарында РНК мен нәруыздар түзілуі азаюы 40 тәуліктің ішінде атрофия дамуына әкеледі. Денервация кезінде тотығулық баяу ет талшықтарында үшкарбон қышқылы оралымының негізгі ферменттерінің, цитохромоксидазаның белсенділігі төмендейді, тотығулық фосфорлану, май қышқылдарының тотығуы қатты әлсірейді. Осымен бірге глюкоза-6-фосфатдегидрогеназа, каталаза ферменттерінің белсенділігі көтеріледі, макроэргиялық (АТФ, КрФ т.б.) түзілуі азаяды, анаэробтық гликолиз артады. Жүйкесізденген ет талшықтарында протеазалардың белсенділігі күшейеді, нәруыздардың ыдырауы артып, олардың деңгейі азаяды. Осыдан актин мен миозиннің құрылымында өзгерістер пайда болады, миозиннің жеңіл тізбектері сұрыпталып азаяды. Жүйкесізденген ет талшықтарында, нәруыздардың жалпы мөлшері азайғанына қарамай, миоциттердің саркоплазмасының шеттерінде бос рибосомалар мен түйіршіктелмеген торшалар жинақталады. Олар Гөлдж аппаратынан көптеген холинорецепторлардың синапстардан тыс аумақтарға тасымалдануына қолайлы жағдай жасайды. Содан холинорецепторлардың бұндай синапстан тыс тығыз орналасуы қанмен айналып жүрген ацетилхолинге ет талшықтарының сезімталдығын көтереді.
Жүйкесізденген ет талшықтарының атрофиясы жүйке талшықтары үзілгеннен кейін 100-120 тәулік аралығында дамиды. Бұл кезде атрофия дамуымен қатар бұлшықетте нөкер-жасушалардың көлемі ұлғаяды. Олар 2-3 аптадан кейін жетілмеген ет талшықтарын қалыптастырады.
Егер бұлшықеттің денервациясы жұлынның тек алдыңғы түбіршегі оқшауланып бүлінуінен болса, онда жүйкелік серпіндерді мыйға бағыттайтын сезімтал афференттік жүйе сақталып қалады және ол өзінің функциялық қасиетін өзгертеді. Денервациялық синдром дамуына қарай сезімтал мембрананың Na+ иондарына өткізгіштігі артады, созып керуге ет талшықтарының сезімталдығы көтеріледі. Сонымен қатар жүйкесізденген ет талшықтарында сезімтал жүйке аяқшаларының өз бетінше электр белсенділігі күшейеді.
Жүйкесізденген бұлшықеттің қайта жүйкеленуі қимылдық жүйке талшықтарының өз бетінше өсуінен немесе хирургиялық әдіспен үзілген жүйке талшығын жалғаудан кейін байқалады. Тікелей қайта жүйкелену үзілген жүйкенің орталық кесіндісінен қимылдық жүйке талшықтарының өсуі арқылы қамтамасыз етіледі. Бұл үрдіс бірнеше сатыда өтеді. Бірінші сатысында жүйкелік-еттік синапстарда алдымен синапс алды, артынан синапс соңы құрылымдар бүлінеді, бүліну өнімдері фагоцитоз жолымен жойылады және жүйкесізденген бұлшықетті қоректендіруге қажетті қанайналым азаяды. Осындай жағдайды Уоллер дегенерациясы дейді. Екінші сатысында жүйкесізденген ет талшықтарынан және шванн жасушаларынан жүйкелердің өсу факторы (ЖӨФ) және молекулалардың адгезиялық факторы бөлініп шығады. Олар үзілген жүйкенің орталық ұшына өсуді сергітетін әсер етеді. Жүйкесізденген бұлшықетте жүйке үзілгеннен кейін 10-12 тәуліктен соң ЖӨФ өндірілуі қатты көтеріледі. Бұл фактор жүйке талшықтарының өсу бағытын анықтайтын хемотаксистік әсер етеді. ЖӨФ сезімтал және дербес жүйке нейрондары аксондарының соңғы рецепторларымен байланысады, сорылулық эндоцитоз жолымен аксонға өтеді және кері бағытта аксоплазмалық ағыммен ядроның айналасына ауысады да онда нейрондық нәруыздардың түзілуін реттейді.
Өсіп келе жатқан жүйке аяқшалары молекулалардың агрегациялық факторын босатып шығарады. Ол жаңа синапстық аумақтардың құрылуын арттырады немесе бар синапстардың көлемін ұлғайтады.
Үшінші сатысында өсіп келе жатқан жүйке талшықтары жүйкесізденген бұлшықетке жетеді және оның жасушаларымен түйіседі. Бұл кезде қалпына келе бастаған қимылдық нейронның денесінде нәруыздар түзілуі артады, олардың аксоплазмалық ағыммен жүйкенің өсіп келе жатқан аяқшаларына тасымалдануы күшейеді. Қалпына келе бастаған аксондар ет талшықтарының бұрынғы синапстық аумақтарында 7 тәулік ішінде «тұрпайы» синапстар қалыптастырады. Оларда әлі электр серпіндік белсенділік болмайды. Бұл синапстарда холинорецепторлардың агрегациясы болады. Ет талшықтарында осы рецепторлардың тығыздығы жоғары деңгейге жеткенде мотонейрон аксондарының аяқшаларына аз мөлшерде нейромедиатор шығарылуынан жекелеген әрекеттік потенциал өндірілу мүмкіншілігі пайда болады.
Төртінші сатысында «тұрпайы» синапстардың жетілуі арқылы нақтыланған жақсы жетілген синапс қалыптасады. Бұл кезде қалпына келе бастаған аксонның серпін туындататын белсенділігі қалпына келуі қажет. Ол түйіспе қатпарлары құрылуын арттырады және, түйіспе асты мембрананы қаптайтын, тіректік мембранада ацетилхолинэстераза жиналуын сергітеді. Бұл үрдіс аяқталғаннан кейін бір айға дейін иондық каналдардың ашылу мерзімі қысқарады, көптеген терминалдық аяқшалар жоғалады және жекелеген жүйке аяқшалары сұрыпталып соңғы табақшада тұрақталады. Бұл кезде синапстық саңылау 50 нм шамасында болады және ол болбыр жасушасыртылық затпен толады. Осымен бір мезгілде синапстан тыс аймақтарда сарколемманың қосымшы түйіспелер қалыптастыру қабілеті жоғалады, холинорецепторлар тек жаңадан құрылған соңғы табақшаларда шоғырланады. Нәрленісті жүйкелік қадағалау қалпына келуі қайта жүйкеленген бұлшықетте қоректенуге қажетті қан айналымды қалпына келтіреді. Бірақ бұлшықеттің қайта жүйкеленуі кезінде ет талшықтары мен сіңір рецепторларының саны азаяды.
Шапшаң бұлшықеттің кесілген жүйкесінің орталық ұшын шабан бұлшықеттің кесілген жүйкесінің шеткері ұшымен хирургиялық жолмен жалғағанда немесе кесілген жүйкенің орталық ұштарын басқа бұлшықетке қосқанда қиылысқан қайта жүйкелену пайда болады. Бұл кезде қайта жүйкелену сатылары қалпына келе бастаған жүйке мен жүйкесізденген сәйкес бұлшықеттің қайта жүйкелену сатыларына ұқсас болады. Дегенмен олардан біршама ерекшеліктері байқалады.
Шапшаң бұлшықеттің жүйкесін шабан бұлшықетке жалғағанда түйіспелік байланыс 22 апта аралығында кеш пайда болады. Өйткені бұл кезде шабан бұлшықеттерге тән тропонин, миозин, тропомиозин жаңа бағдарлама бойынша түзілуінің үйлесімі бұзылады. Әр ет талшықтарында шапшаң бұлшықеттерге ғана тән жоғарыда келтірілген жиырылғыштық нәруыздар байқалады. Шапшаң бұлшықеттің жүйкесімен жүйкеленген шабан бұлшықеттің жиырылу күші мен жылдамдығы қатты артады.Сонымен қатар онда гликолиздік белсенділік күшейеді. Ацетилхолинге сезімталдық негізінен соңғы табақшаларда шоғырланады, өз бетінше электр әлеуетін туындату қабілеті жоғалады. Қайта жүйкеленген бұлшықетте ет талшықтары мен сіңір рецепторларының саны азаяды.
Шапшаң бұлшықетке шабан бұлшықеттің жүйкесін жалғағанда да керісінше жағдай байқалады. Шапшаң бұлшықеттерге тән тропонин, миозин, тропомиозин түзілуі бұзылады, шабан бұлшықеттерге тән нәруыздар түзіле бастайды, шапшаң бұлшықет шабан бұлшықеттің қасиетін қабылдап, оның жиырылу күші мен жылдамдығы төмендейді, онда тотығулық ферменттердің белсенділігі көтеріліп, гликолиздің қарқыны азаяды. Сонымен қайта жүйкеленген бұлшықетте шабан бұлшықетке тән сарколемманың ацетилхолинге сезімталдығы пайда болады.
Қозудың жүйкеден бұлшықетке тарауы және жүйкелік нәрленістік ықпалдардың бұзылыстары
Жүйке-бұлшықеттік түйіспе қимылдық аксонның аяқшасынан, синапс саңылауынан және ет тінінің синапс соңылық мембранасынан тұрады. Қимылдық нейронның денесі ядросының айналасында және жүйке аяқшаларының аксоплазмасында холинацетилтрансфераза ферментінің қатысуымен ацетилхолин түзіледі. Оның 80% ға дейіні түйіспелік көпіршіктерге түседі. Бұл көпіршіктер пресинапстық мембрананың алдында жинақталады. Пресинапстық және постсинапстық мембраналардың арасында мөлшермен 60 нм синапстық саңылау болады. Ол жүйке және ет талшықтарының арасын байланыстыратын коллаген тәрізді болбыр затпен толады. Синапс соңы мембранада шынайы холинэстераза ферментінің молекулалары орналасқан. Холинорецепторлар постсинапстық мембрананың қатпарларының шыңдарындағы жасуша қаңқаларына бекітілген. Түйіспелерде өз бетінше және жүйкелік серпіндердің әсерлерінен нейромедиаторлар босап шығады. Өз бетінше нейромедиатордың босауы постсинапстық мембрананың жергілікті тұрақты деполяризациясын және сол арқылы бұлшықеттің қалыпты межеқуатын қамтамасыз етеді. Нейромедиатордың бірмезгілдендірілген квантты босауы соңғы табақшаларда потенциал туындатады, артынан,пресинапстық аймаққа аксонның бойымен әрекеттік потенциал түсуіне жауап ретінде, ет талшықтарында әрекеттік потенциал пайда болады. Жүйке аяқшаларының деполяризациясы электр серпіндердің әсерінен болады және ол потенциал-тәуелді Са2+ өзекшелерін ашып, Са2+ иондарының жасушалар ішіне енуін қамтамасыз етеді. Жүйке аяқшаларының реполяризациясы кезінде Са2+ иондары жасушалар сыртына шығарылады. Жүйке аяқшаларының ішіне енген Са2+ иондары, кальмодулинмен байланысып, синапстық саңылауға ацетилхолин босап шығуына ықпал етеді. Ацетилхолиннің бір молекуласы холинорецептормен байланысуы иондық каналды ашып, Na+ иондарының жасушаларға қысқа мерзімде енуін қамтамасыз етеді. Осыдан пайда болған жергілікті катион ағымдарының жинағы нәтижесінде соңғы табақшаның потенциалы, артынан ет талшығының әрекеттік потенциалы туындайды.
Жүйке-бұлшықет синапсының қалыпты түрткілену жиілігі шамамен минутына 20 серпінге дейін болады.
Қозудың жүйкеден бұлшықетке тарауы және жүйкелік нәрленістік ықпалдардың бұзылыстары:
• нейромедиатордың түзілуі ауытқуларынан,
• нейромедиатордың аксонның бойымен майда көпіршіктерге тасымалдануы бұзылыстарынан,
• нейромедиатордың қимылдық нейрондардың аяқшаларында жинақталуы ақауларынан,
• синапстық саңылауларда ацетилхолинэстеразаның қатысуымен ацетилхолиннің ыдыратылуы баяулауынан,
• постсинапстық мембранада холинорецепторлардың түзілуі, ыдыратылуы және жинақталуы бұзылыстарынан,
• жүйкелік серпіндерге жауап ретінде холинорецепторларда иондық өзекшелердің ашылу мерзімі мен тәртібі ауытқуларынан (баяу каналдық синдром) – байқалады.
Ацетилхолиннің түзілуі мен оның қимылдық жүйкелердің аяқшаларында жинақталуының ақаулары
Нейромедиаторлардың түзілуі, аксонның бойымен тасымалдану








