 2014-01-31
2014-01-31 1074
1074Различными методами показано, что в период, предшествующий делению клеток, количество в них ДНК удваивается. При этом синтез ДНК осуществляется таким образом, что и в дочерних, и в материнских клетках содержатся совершенно идентичные молекулы ДНК. Это означает, что в процессе синтеза реализуется механизм воспроизведения структуры ДНК, который обеспечивает передачу наследственных свойств в ходе размножения клеток. Механизм образования в клетках организмов новых молекул ДНК, которые идентичны по генетическим свойствам уже имеющимся молекулам, называют репликацией ДНК.
Первые представления о механизме удвоения ДНК вытекали из модели строения молекул ДНК Д. Уотсона и Ф. Крика, которая представляет собой двойную спираль, состоящую из двух комплементарных полинуклеотидных цепей. Если такие цепи разъединить и использовать в качестве матрицы (шаблона) для синтеза новой цепи ДНК, то в процессе синтеза будет выстраиваться полинуклеотидная цепь, которая по всем нуклеотидам комплементарна исходной цепи ДНК.
Таким образом, каждая из двух разъединённых цепей ДНК даёт начало новой двухцепочечной структуре, которая по последовательности в ней нуклеотидных остатков не отличается от исходной молекулы ДНК, используемой в качестве матрицы при синтезе новой молекулы ДНК. Принцип изложенного выше механизма удвоения ДНК может быть представлен в виде следующей схемы:
|
Экспериментальное подтверждение механизма репликации ДНК было получено в 1958 г. М. Мезелсоном и Ф. Сталем, которые выполнили эксперименты с использованием тяжёлого изотопа азота 15N. Путём длительного выращивания кишечной палочки (E. сoli) на среде, содержащей в качестве источника азота 15NH4Cl, они получили клетки, в которых молекулы ДНК были полностью насыщены 15N. Затем клетки перенесли в среду с источником азота в виде 14NH4Cl и стали отбирать пробы после появления каждой новой генерации клеток.
Из каждой пробы выделяли ДНК и подвергали ультрацентрифугированию в градиенте плотности хлористого цезия. В ходе опытов было определено, что в первом поколении клеток после их высаживания на среду с 14N в составе ДНК только половина азота была представлена в виде 15N, а вторую половину составлял изотоп 14N. Такое распределение изотопов азота удалось определить путём разделения цепей ДНК при нагревании, в одной из цепей содержался изотоп азота 15N, а в другой – 14N. Полученные результаты свидетельствовали о том, что в ходе синтеза образуется гибридная молекула ДНК, в которой одна из цепей, содержащая 15N, переходит от родительской молекулы, а вторая синтезируется заново с использованием азота из питательной среды в виде 14N.
В следующем акте деления клеток гибридная молекула ДНК разделялась на две цепи и на каждой из цепей синтезировалась новая полинуклеотидная цепь, используя в качестве источника азота только изотопы 14N. (рис. 45). Поэтому каждая гибридная молекула ДНК в ходе репликации давала начало одной новой гибридной молекуле и другой уже не гибридной молекуле, содержащей комплементарные цепи ДНК и имеющей атомы азота только в виде 14N. В каждом последующем поколении происходило уменьшение доли гибридных молекул в два раза.
Синтез ДНК катализируют ферменты ДНК-нуклеотидилтрансфе-разы или более кратко ДНК-полимеразы, имеющие в активном центре катионы Zn2+. ДНК-полимеразы осуществляют синтез ДНК на одноцепочечной матрице, если на ней уже содержится небольшой олигонуклеотидный фрагмент, комплементарный нуклеотидным остаткам матрицы. Это так называемая «затравка», у которой на 3'-конце имеется свободная гидроксильная группа. Субстратами для построения синтезируемой полинуклеотидной цепи ДНК служат дезоксирибонуклеозидтрифосфаты, то есть дАТФ, дГТФ, дЦТФ, дТТФ. Порядок присоединения фосфорилированных нуклеотидных остатков определяется их комплементарным взаимодействием с соответствующими нуклеотидными остатками на ДНК-матрице, с которыми они образуют водородные связи.
В ходе реакции под действием фермента осуществляется гидролиз макроэргической связи дезоксинуклеозидтрифосфата и высвобождение пирофосфата, а нуклеотидный остаток в виде дезоксинуклеотидмонофосфата присоединяется к 3'-ОН «затравки». И так последовательно происходит наращивание синтезируемой полинуклеотидной цепи ДНК в направлении от 5'-конца к 3'-концу. Вновь синтезируемая цепь ДНК комплементарна и антипаралельна полинуклеотидной цепи матрицы, что отвечает принципам построения двойной спирали ДНК. Реакция, катализируемая ДНК-полимеразой, может быть представлена в виде следующей схемы:
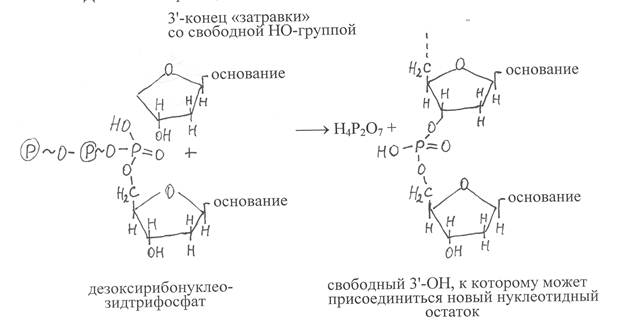
Образующийся в указанной реакции пирофосфат расщепляется пирофосфатазой, что предотвращает ход обратной реакции, вызывающей распад полинуклеотидов ДНК.
ДНК-полимеразы осуществляют синтез полинуклеотидных цепей ДНК с очень высокой скоростью. Скорость этого процесса в клетках бактерий составляет около 1000 нуклеотидных остатков за 1 секунду. В клетках высших организмов – несколько десятков нуклеотидных остатков за 1 секунду (около 100). Однако даже при такой высокой скорости синтеза фермент действует очень точно и вероятность присоединения неправильного нуклеотидного остатка обычно не превышает 10-6 – 10-8, то есть ошибка в синтезе ДНК составляет один раз на десятки миллионов нуклеотидных остатков.
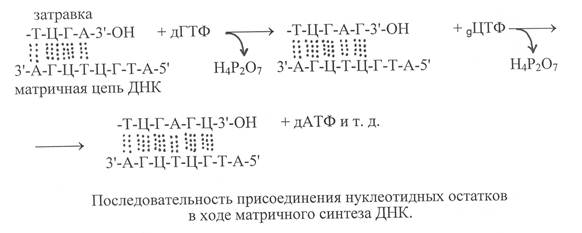 |
Как было указано выше, ДНК-полимеразы присоединяют нуклеотиды в ходе синтеза ДНК к 3'-концам олигонуклеотида, называемого «затравкой», комплементарно присоединённого к матрице. Однако эти ферменты не способны инициировать синтез ДНК на одноцепочечной матрице. Поэтому образование олигонуклеотида «затравки» катализирует другой фермент, который назвали ДНК-праймазой. ДНК-праймаза катализирует присоединение к матричной цепи ДНК нескольких остатков рибонуклеотидов, формирующих короткую «затравочную» цепь РНК, которая комплементарна матричной цепи и образует с ней двойную спираль. Именно к 3'-концу РНК-затравки происходит далее присоединение остатков дезоксирибонуклеотидов, катализируемое ДНК-полимеразами. После того, как синтез новой ДНК-цепи заканчивается, РНК-затравки удаляются специальным ферментом, который проявляет 5'-экзонуклеазную активность, то есть отщепляет от полинуклеотидной цепи 5'-концевые нуклеотидные остатки.
В клетках бактерий выявлены три вида ДНК-полимераз – ДНК-полимеразы I, II, и III. ДНК-полимераза I представляет собой белок с молекулярной массой 102000, обладающий тремя видами каталитической активности. Он способен удалять неправильно присоединённые или изменённые в результате мутации нуклеотидные остатки за счёт проявления 3'-экзонуклеазной активности и присоединять вместо них правильные нуклеотидные остатки, комплементарные матричной ДНК. Кроме того, за счёт 5'-экзонуклеазной активности этот фермент катализирует удаление РНК-затравки и достраивание вместо неё полинуклеотидной цепи ДНК.
ДНК-полимераза II имеет молекулярную массу около 120000 и обладает как полимеразной, так и 3'-экзонуклеазной активностью. Она присоединяется к двуцепочечной ДНК и катализирует на одной из цепей ДНК образование фрагментов длиной в несколько десятков нуклеотидов. Важную роль этот фермент играет в процессах репарации ДНК, направленных на удаление в ДНК изменённых под воздействием мутагенов нуклеотидных остатков.
ДНК-полимераза III – главный фермент, катализирующий репликацию ДНК. Он состоит из семи разных субъединиц, образующих олигомерную молекулу с молекулярной массой около 500000. Ферментный белок присоединяется к двуцепочечной затравке и катализирует далее синтез новой цепи ДНК на ДНК-матрице. На активирование ДНК-полимеразы III в процессе присоединения к матрице затрачивается энергия, которая обеспечивается в результате гидролиза макроэргической связи АТФ с образованием АДФ и фосфорной кислоты.
В клетках высших организмов имеются аналогичные ферменты, обладающие ДНК-полимеразной активностью. Основной фермент репликации ДНК в ядрах клеток – ДНК-полимеразе a, в составе которого содержится также полипептидная субъединица ДНК-праймазы, способная катализировать синтез РНК-затравки. ДНК-полимераза b, как и ДНК-полимераза II бактерий, участвует в репарации ДНК. ДНК-полимераза ¡ катализирует синтез ДНК в митохондриях. Аналогичный фермент найден в хлоропластах растений.
У бактерий репликация ДНК начинается в определённом участке хромосомы, который сокращённо обозначают ori (от английского слова origin – начало). Этот участок хромосомы имеет специфическую последовательность нуклеотидов, узнаваемую одним из белков, участвующим в инициации синтеза ДНК. Его называют инициаторным белком. При этом может быть однонаправленная репликация, когда ферменты репликации движутся в одном направлении. Возможен также двунаправленный синтез ДНК, когда репликация ДНК от участка хромосомы, обозначаемого ori, осуществляется в двух противоположных направлениях. В хромосомах высших организмов репликация начинается одновременно на многих участках, расстояние между которыми составляет порядка 100000 нуклеотидных остатков.
После того, как инициаторный белок связывается с участком ДНК ori, к этому участку присоединяется в определённой последовательности комплекс из шести белков, главным из которых является хеликаза. Хеликаза – АТФ-зависимый фермент, использующий энергию гидролиза АТФ для разъединения полинуклеотидных цепей двойной спирали ДНК. Ферментный белок хеликазы, присоединившись к одной из цепей ДНК, движется вдоль этой цепи и расплетает перед собой двойную спираль ДНК, образуя репликативную вилку, которая включает участок двойной спирали ДНК и две отходящие от него одноцепочечные ветви (рис. 46).
Самопроизвольное объединение в двойную спираль образовавшихся под действием хеликазы одноцепочечных участков ДНК предотвращают специальные белки, которые имеют избирательное сродство к разъединённым полинуклеотидным цепям ДНК. Такие белки получили название SSB-белков (от английского single strand binding proteins). SSB-белки, вплотную прилегая друг к другу, связывают одноцепочечные участки ДНК и делают их более реакционноспособными для взаимодействия с ферментом ДНК-полимеразой.
В результате расплетания хеликазой двойной спирали ДНК возникает её вращательное движение впереди репликативной вилки. Однако в связи с тем, что хромосома связана и закреплена клеточными мембранами, вращение двуспиральной ДНК приводит к образованию дополнительных витков и сверхскрученных узлов на пути продвижения репликативной вилки, что могло бы в дальнейшем остановить процесс репликации ДНК. Но этого не происходит вследствие того, что на участке сверхспирализации впереди по движению репликативной вилки действует фермент топоизомераза, образующий разрыв фосфоэфирной связи в одной из цепей двуспиральной ДНК. За счёт такого действия топоизомеразы обеспечивается свободное вращение оставшейся неразорванной одноцепочечной нити ДНК вокруг её υ-связи, что приводит к снятию сверхспирализации ДНК на участке продвижения репликативной вилки и раскручиванию образовавшихся узлов, вызванных действием фермента хеликазы. После раскручивания дополнительных витков сверхспирализации топоизомера восстанавливает разорванную фосфодиэфирную связь и таким образом сохраняет структурную целостность двойной спирали ДНК, которая участвует в процессе репликации.
В результате действия хеликазы и SSB-белков образуются две разъединённые цепи ДНК, имеющие противоположную направленность. Одна из них начинается от 3'-конца, другая – от 5'-конца. На цепи ДНК, начинающейся с 3'-конца, после образования РНК-затравки возможен непрерывный синтез новой цепи ДНК под действием фермента ДНК-полимеразы, способного присоединять к 3'-ОН растущей цепи новые нуклеотидные остатки. Однако на цепи ДНК, начинающейся с 5'-конца, такой синтез новой цепи ДНК невозможен. Он должен происходить в обратном направлении после образования одноцепочечного участка ДНК определённой длины, вследствие чего синтез ДНК на одноцепочечной ветви репликативной вилки, начинающейся с 5'-конца, происходит медленней, чем на другой одноцепочечной ветви, начинающейся с 3'-конца. В связи с этим одну из синтезирующихся цепей ДНК называют ведущей, а другую цепь, синтез которой происходит медленней, – отстающей.
В опытах Р. Оказаки было показано, что отстающая цепь ДНК синтезируется в виде коротких фрагментов, называемых фрагментами Оказаки. Каждый такой фрагмент начинается с синтеза РНК-затравки с участием ДНК-праймазы, а далее действует фермент ДНК-полимераза III (у бактерий) или ДНК-полимераза a (у высших оорганизмов), который уже синтезирует ДНК-цепь фрагмента Оказаки, доводя её до следующей РНК-затравки. После этого в действие включается фермент ДНК- полимераза, обладающая 5'-экзонуклазной активностью (у бактерий это ДНК-полимераза I), которая удаляет впереди себя РНК-затравку и вместо неё достраивает цепь ДНК до следующего фрагмента Оказаки. На стыке двух фрагментов Оказаки имеется с одной стороны 3'-ОН, а с другой 5'-фосфатная группа, между которыми под действием фермента ДНК-лигазы образуется фосфодиэфирная связь, соединяющая фрагменты Оказаки в единую синтезируемую цепь ДНК.
Линейные размеры фрагментов Оказаки зависят от вида организмов; у бактерий они составляют в длину до 1000 нуклеотидных остатков, у высших организмов – около 100.
Наиболее хорошо процесс репликации ДНК изучен у бактерий Е. cоli (рис. 47). В клетках E. coli формирование репликативной вилки инициируют два вида хеликаз. Одна из них образует комплекс с ферментным белком праймазы, который получил название праймосомы. Праймосома, расплетая двуцепочечную ДНК, движется по отстающей её цепи в направлении от 5'-конца к 3'-концу и синтезирует на этой цепи РНК-затравки для фрагментов Оказаки. Вторая хеликаза (вспомогательная) движется по ведущей цепи ДНК от её 3'-конца к 5'-концу. Впереди по движению репликативной вилки действует фермент ДНК-гираза, снимающий витки сверхспирализации двойной спирали ДНК, которые возникают в результате работы хеликаз. Образовавшиеся в репликативной вилке одноцепочечные участки ДНК связываются SSB-белками и сразу же становятся объектами действия для фермента ДНК-полимеразы III.
Одна молекула ДНК-полимеразы III движется по одной из ветвей репликативной вилки и синтезирует ведущую цепь ДНК. Другая молекула этого же фермента движется по другой одноцепочечной ветви и катализирует образование фрагментов Оказаки, достраивая полинуклеотидные участки от одной затравки до другой. После этого фермент ДНК-полимераза I удаляет РНК-затравки и вместо них достраивает фрагменты Оказаки нуклеотидными остатками ДНК. На конечном этапе синтеза отстающей полинуклеотидной цепи ДНК фермент ДНК-лигаза соединяет фрагменты Оказаки фосфодиэфирными связями в единую цепь.
В основных чертах механизм репликации ДНК у разных организмов имеет много общего, хотя у высших организмов по сравнению с низшими имеются определённые особенности. Многие из них уже показаны в данной главе. Следует добавить ещё одно отличие механизма репликации ДНК у высших организмов по сравнению с низшими. Оно связано со строением хромосом. У высших организмов хромосомы линейны и у них на 5'-конце отстающей цепи ДНК остаётся недореплицированный участок в связи с неспособностью ДНК-полимераз инициировать синтез ДНК на одноцепочечной матрице. Для того, чтобы репликация ДНК проходила полностью, у высших организмов имеются удлинения на концах хромосом из повторяющихся последовательностей нуклеотидов, которые образуют изгиб с разворотом цепи на 1800, так что свободных концов у хромосомы фактически нет. Присоединение к концам хромосом повторяющихся последовательностей нуклеотидов катализирует специальная трансфераза. Длина концевых фрагментов хромосом контролируется регуляторной системой клетки.
Как было указано ранее, репликация ДНК в клетках организмов осуществляется с очень высокой точностью, однако с небольшой частотой ошибки всё-таки происходят и в структуру новосинтезируемой цепи ДНК включаются нуклеотидные остатки, которые не комплементарны матрице. Такие ошибки репликации исправляют ферменты, относящиеся к системе репарации ДНК. Процесс репарации ДНК – это исправление ошибок репликации ДНК и устранение изменённых нуклеотидных остатков, которые возникают под воздействием мутагенных факторов – радиации, ультрафиолетового излучения, алкилирующих химических веществ, дезаминирования и потери оснований в нуклеотидных остатках.
Функционирование системы репарации ДНК обеспечивает исключение из её структуры неправильно присоединённых или изменённых под действием мутагенных факторов нуклеотидных остатков и таким образом устойчиво поддерживает стабильность содержащейся в молекулах ДНК генетической информации. Однако с небольшой частотой иногда такие ошибки возникают и дают начало при репликации изменённым молекулам ДНК. Появление в результате ошибок репликации или действия мутагенных факторов молекул ДНК с изменённым составом нуклеотидных остатков получило название генетических мутаций.

