 2018-03-09
2018-03-09 1034
1034
Желчные кислоты по своему строению близки к холестерину и являются производными холановой кислоты.
Желчные кислоты (холевая, дезоксихолевая, лихотолевая) входят в состав желчи как в чистом виде, так и в виде парных соединений с гликоколом (глицином) и таурином (с которыми они соединяются посредством пептидной ковалентной связи).

Для открытия желчных кислот используют их способность давать красное окрашивание с оксиметилфурфуролом (реакция Петтенкофера). Оксиметилфурфурол образуется при реакции фруктозы с концентрированной соляной или серной кислотой.
В сухую пробирку наливают 10 капель разведенной желчи, добавляют 1-2 капли раствора сахарозы (или фруктозы) и, наклонив пробирку, осторожно (по стенке) наливают равный объем концентрированной серной кислоты. На границе слоев образуется пурпурное кольцо, которое затем принимает красно-фиолетовое окрашивание.
Контрольные вопросы к отчету по теме «Липиды»
1. Что такое липиды?
2. По какому признаку классифицируются липиды?
|
|
|
3. Напишите формулы важнейших представителей всех классов липидов и назовите их.
4. Какими константами характеризуются свойства жиров?
5. Что такое кислотное число жира?
6. Что такое иодное число жира?
7. Что такое число омыления?
8. Какие ферменты производят гидролитическое расщепление жиров в органах и тканях?
9. Какова роль желчи в переваривании и всасывании жиров?
10. Напишите реакции гидролиза жира и фосфатидилхолина.
11. Как глицерин окисляется в тканях?
12. Какова роль карнитина в окислении жирных кислот?
13. Напишите реакции b-окисления капроновой кислоты.
14. Где происходит окисление жирных кислот?
15. Как окисляются ненасыщенные жирные кислоты?
16. Каков баланс энергии окисления жирных кислот?
17. Какие ткани являются жировыми депо?
18. Что такое кетоновые тела? Как они образуются?
19. Как происходит биосинтез жирных кислот? Укажите роль АПБ.
20. Где происходит биосинтез жирных кислот?
21. Как идет синтез триацилглицеринов?
22. Запишите реакции образования глицерофосфолипидов.
23. Напишите уравнения реакций, протекающих по следующей схеме:

Напишите промежуточные и конечные продукты реакций.
Занятие 7. Зачет по всем темам и защита реферативной работы
БИОСИНТЕЗ БЕЛКА
ДНК – молекула наследственности у всех прокариотических и эукариотических организмов (у вирусов роль генетического материала выполняют либо ДНК, либо РНК). Любая клеточная ДНК представляет собой две очень длинные спиральные полинуклеотидные цепи, закрученные вокруг общей оси. Цепи антипараллельны. Сахарофосфатный остов каждой цепи лежит снаружи двойной спирали, а пуриновые и пиримидиновые основания – внутри. Две цепи удерживаются вместе водородными связями между парами оснований (аденин образует две водородные связи с тимином (А-Т); гуанин – три водородные связи с цитозином (G-C). Цепи двойной спирали взаимно комплементарны. Генетическая информация закодирована в последовательности оснований вдоль цепи.
|
|
|
Функция ДНК состоит в том, что она хранит запас генетической информации, необходимой для кодирования структуры всех белков и всех РНК каждого вида организмов, регулирует во времени и пространстве биосинтез компонентов клеток и тканей, определяет деятельность организма в течение его жизненного цикла и обеспечивает индивидуальность данного организма.
Гены – это участки ДНК, которые кодируют полипептидные цепи и РНК. Часть генов кодирует, например, разные виды тРНК, другие гены отвечают за синтез различных рРНК. Гены, кодирующие полипептиды и РНК, называются структурными генами, так как они определяют структуру определенного продукта гена – фермента или стабильной РНК. В ДНК содержатся и другие участки или последовательности, которые выполняют исключительно регуляторную функцию. Некоторые из этих регуляторных участков представляют собой сигналы, обозначающие начало и конец структурных генов; другие принимают участие в запуске или прекращении транскрипции структурных генов. Таким образом, наряду со структурными генами, хромосома содержит также регуляторные последовательности.
В одной хромосоме сосредоточено большое количество генов. В единственной хромосоме бактерии кишечной палочки (E.Coli) содержится более 3000 генов (возможно 5000).
С помощью различных генетических подходов была установлена последовательность расположения многих генов в хромосомах вирусов и бактерий.
Поток генетической информации в нормальных клетках происходит в направлении: ДНК→РНК→ белок. Синтез РНК по ДНК-матрице называется транскрипцией, а синтез белка по РНК-матрице – трансляцией.
Первый этап – репликация, т. е. копирование родительской ДНК с образованием дочерних молекул ДНК, нуклеотидная последовательность которых комплементарна нуклеотидной последовательности родительской ДНК и однозначно определяется ею.
Таким образом, центральная догма молекулярной генетики, показывающая перемещение генетической информации в ходе трех фундаментальных процессов: репликации, транскрипции и трансляции -может быть выражена схемой:

РЕПЛИКАЦИЯ
Репликацией называют процесс удвоения ДНК. Принципиальный механизм его определяется строением двухспиральной молекулы ДНК (рис.1). В результате репликации образуются две молекулы ДНК, представляющие собой точные копии исходной молекулы. Каждая из вновь образующихся молекул содержит одну цепь исходной ДНК и одну вновь синтезируемую цепь. Иными словами, репликация полуконсервативна – половина родительской молекулы сохраняется в дочерней.
Этот процесс даже в простейших случаях реализуется путем сочетания многих сложных механизмов, в которые вовлечены многочисленные ферменты и регуляторные белки. Лучше всего процессы репликации изучены для наиболее простейших систем – бактерий, бактериофагов и плазмид.
Расплетание родительской ДНК и синтез новой ДНК происходит в репликационной вилке. Репликационная вилка – участок ДНК, где происходит одновременное расплетание и синтез новых цепей ДНК. Репликация начинается в строго определенном месте и продолжается последовательно в двух направлениях с примерно одинаковой скоростью.
Исходя из скорости движения репликативной вилки E.Coli, можно заключить, что при 37˚С новая ДНК синтезируется со скоростью свыше 4500 нуклеотидных остатков в минуту в расчете на одну вилку. Поскольку на каждый виток двойной спирали приходится приблизительно 10 пар оснований, скорость расплетания родительской ДНК в репликационной вилке у клеток E.Coli составляет 4500 об./мин., что превышает скорость вращения вала в двигателе автомобиля, мчавшегося со скоростью 110 км/час.
|
|
|
Участок начала репликации представляет собой нуклеотидную последовательность длиной 100-200 пар оснований. Эта последовательность узнается специфическими клеточными белками, которые начинают в этом месте цикл репликации. Именно этот процесс инициации репликации находится под контролем клеточной регуляции.
Как теперь известно, репликация эукариотических ДНК начинается одновременно во многих точках (около 1000). Из каждой точки одновременно в противоположные стороны движутся две репликативные вилки (рис. 2).

 Благодаря этому репликация целой эукариотической хромосомы может завершаться даже быстрее, чем репликация бактериальной хромосомы.
Благодаря этому репликация целой эукариотической хромосомы может завершаться даже быстрее, чем репликация бактериальной хромосомы.
Ключевую роль в процессе репликации играют реплицирующие ДНК-полимеразы, которые осуществляют матричный синтез ДНК из дезоксинуклеозидтрифосфатов. Фермент синтезирует нить ДНК, комплементарную родительской нити (называемой матрицей), последовательно присоединяя к 3´-концу (растущей цепи) мононуклеотидные звенья, комплементарные звеньям матрицы:
(ДНК)n остатков + dNTP → (ДНК)n+1 + PPi.
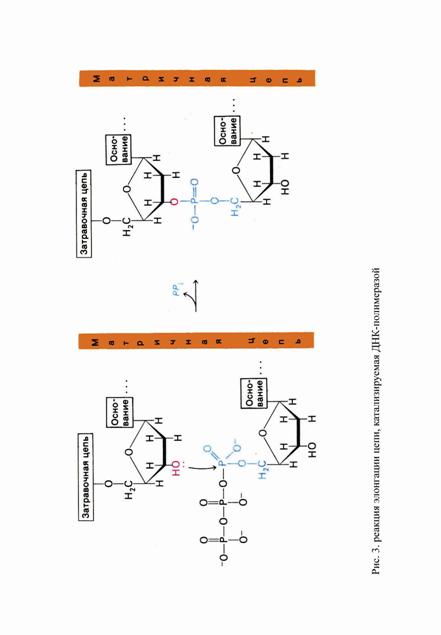
Последующий гидролиз пирофосфата обеспечивает дальнейшую полимеризацию. Элонгация цепи (рост цепи) ДНК происходит в направлении 5´-3´. За 1 сек молекула ДНК-полимеразы присоединяет 10 нуклеотидов. Полимеризация процессивна, т. е. фермент присоединяет много нуклеотидов, оставаясь связанным с одной матрицей. При этом ДНК-полимераза катализирует нуклеофильную атаку 3´-ОН группы концевого нуклеотида растущей цепи на α-фосфатную группу дезоксинуклеозидтрифосфата, отбираемого ферментом на основе его комплементарности соответствующему звену матрицы. В результате отщепляются пирофосфаты и образуется фосфодиэфирная связь. Растущая цепь удлиняется на одно звено, и процесс повторяется с новым дезоксинуклеозидтрифосфатом.
|
|
|
Для репликации необходима не только ДНК-полимераза. В этом процессе участвует больше двадцати различных ферментов и белков, каждый из которых выполняет определенную функцию.
Репликация состоит из большого количества последовательных этапов, в которые включают узнавание точки начала репликации, расщепление родительского дуплекса, удержание его цепей на достаточном расстоянии друг от друга, инициацию синтеза новых дочерних цепей, их элонгацию, закручивание цепей в спирали и, наконец, терминацию репликации. Все эти процессы протекают с очень высокой скоростью и исключительной точностью. Весь этот комплекс называют ДНК-репликазной системой, или реплисомой.
Клетки E.Coli содержат три различные ДНК-полимеразы:
| Катализируемая активность | ДНК-полимеразы бактерии E. Coli | ||
| I | II | III | |
| 5´-3´ полимеразная | + | + | + |
| 5´-3´ экзонуклеазная | + | - | + |
| 3´-5´ экзонуклеазная | + | + | + |
| М.М. | 10900 | 120000 | 400000 |
| Количество молекул в одной клетке | 400 | 10 | |
| Активность (число присоединений за минуту в расчете на одну молекулу) 37˚С | 600 | 30 | 9000 |
Полимеразы кроме полимеразной активности имеют 5´-3´ и 3´-5´ экзонуклеазную активности, т. е. они могут отщеплять концевые нуклеотиды с любого конца цепи ДНК.
Для того чтобы ДНК-полимеразы могли начать синтез, необходимо существование уже готового фрагмента ДНК или РНК, комплементарного матрице и содержащего свободную 3´-ОН-группу. Этот фрагмент называют «затравкой». Инициация репликации начинается с расплетания нитей родительской ДНК и синтеза короткого РНК-транскрипта, который служит праймером («затравкой») для синтеза ДНК. Праймер синтезируется специальными ферментами - праймазами. В E.Coli праймаза активируется примосомой – мультиферментным белковым комплексом (7 - субъединиц).
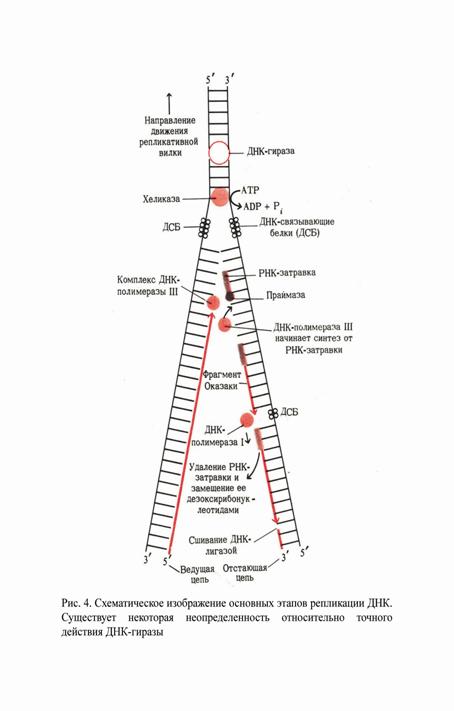
После инициации начинается продвижение репликативных вилок – элонгация. Одна из цепей вновь синтезируемой ДНК удлиняется в том же направлении, в котором движется репликативная вилка, причем синтез осуществляется непрерывно. Эту цепь ДНК называют инициирующей (ведущей). Другая цепь – запаздывающая (отстающая) - синтезируется короткими фрагментами Оказаки. Синтез каждого фрагмента инициируется вблизи начала репликационной вилки и продолжается в противоположную от нее сторону до тех пор, пока 3´-конец вновь синтезируемой ДНК не достигает 5´-конца предыдущего фрагмента Оказаки. Инициация синтеза этих фрагментов осуществляется праймазой, которая в качестве «затравки» синтезирует короткие фрагменты РНК, далее они удлиняются ДНК-полимеразой III. Длина фрагмента Оказаки равна примерно 1000 нуклеотидов. В репликативной вилке работает также еще два белка. Один из них (ДСБ´ белок) специфически связывает одноцепочечные ДНК, облегчая расплетание двойной спирали и одновременно защищая одноцепочечные участки от действия нуклеаз. Другой – ДНК-расплетающий белок (хеликаза) _ движется вдоль двойной цепи, расплетая ее при одновременном гидролизе АТФ. Еще один фермент – ДНК-гираза (ДНК-топоизомераза II) предотвращает накопление супервитков, вызываемое расплетением двойной спирали, образуя в циклически ковалентно замкнутой ДНК одноцепочечные разрывы и затем снова «сшивая» их.
Фермент ДНК-полимераза I удаляет РНК-затравку и достраивает фрагменты, а ДНК-лигаза соединяет между собой соседние фрагменты Оказаки.








