 2014-02-03
2014-02-03 1728
1728Рис.7. Перенос фосфата в матрикс энергизованной митохондрии.
Рис.6. Перенос кальция в митохондрии
При энергизации митохондрии на мембране создается разность Dj (внутри – минус). В мембране митохондрий имеется переносчик ионов кальция, через который они могут проходить вместе со своим зарядом под действием электрического поля (электрофоретически). Как видно из уравнения 2, при этом на мембране снижается Dj и возрастает DpH.
С другой стороны, между матриксом и окружающей средой в энергизованной митохондрии имеется разность pH (более щелочная среда внутри). Это приводит к диффузии внутрь митохондрий ортофосфата, для которого в мембране имеется переносчик, работающий по механизму антипорта: ион HPO3- обменивается на гидроксил OH- (рис. 7). В водном растворе это эквивалентно симпорту (совместному переносу) H+ и HPO3-. При таком переносе не происходит переноса заряда через мембрану, и поэтому перенос не зависит от мембранного потенциала. Однако разность концентрации H+ способствует переносу фосфата в матрикс по закону диффузии для ионов гидроксила,
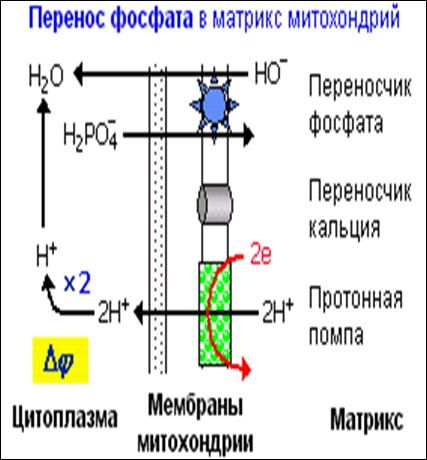
В мембране митохондрий имеется переносчик фосфата, который работает по принципу ионообмена: перенос H2PO4- в матрикс сопровождается переносом HO- - в противоположную сторону (антипорт). Это означает, что при переносе фосфата в матриксе повышается концентрация протонов.
При энергизации митохондрии на мембране создается разность pH (внутри - щелочная среда). Это становится движущей силой для диффузии гидроксильных ионов наружу, но такая диффузия возможна только одновременно с переносом внутрь H2PO4- через переносчик фосфата. Поэтому при переносе фосфата на мембране снижается DpH и возрастает Dj.
которых в матриксе больше, чем снаружи. Таким образом, ионы кальция накапливаются в матриксе электрофоретически, при этом мембранный потенциал снимается, а разность pH на мембране возрастает. Фосфат переносится в матрикс в конечном счете в электронейтральной форме, при этом снижается разность pH, а мембранный потенциал растет. Если в среде присутствуют одновременно и ионы кальция и ортофосфат, происходит накопление обоих в матриксе, при этом снимается как мембранный потенциал, так и DpH. Ничто более не препятствует свободному прохождению ионов водорода через мембрану в точках сопряжения, и дыхательная цепь работает на полную мощность. Из теории Митчелла следует, что прекращение синтеза АТФ митохондриями (или подавление транспорта кальция и фосфата в матрикс) может произойти не только в результате повреждения дыхательной цепи (и снижения скорости дыхания, т.е. окисления субстратов кислородом), но и в случае нарушения барьерных свойств внутренней мембраны. В последнем случае происходит утечка ионов через поврежденную мембрану и падение разности потенциалов на мембране, необходимой для работы АТФ-синтазы, транспорта фосфата и ионов кальция.
Изучение функций митохондрий и их нарушения производится после выделения этих органелл из ткани; при этом важно не повредить митохондрии в ходе самого выделения. Один из способов изучения функции митохондрий - измерение скорости потребления кислорода суспензией органелл в различных функциональных состояниях методом полярографии. На рис. 8 приведена схематизированная запись изменения концентрации кислорода в суспензии в ходе инкубации изолированных митохондрий.

