 2014-02-17
2014-02-17 1416
1416Банки генов (геномная библиотека)
Клонирование генов
Выделение генов
Донорная система
Как известно, геном ядра клеток высших растений содержит большое количество ДНК. Выделение фрагмента ДНК с известной нуклеотидной последовательностью, составляющий определённый ген, из десятка миллионов подобных - представляет собой непростую задачу.
Существуют несколько способов решения:
1. Химический синтез гена.
Возможен при наличии подробной информации о нуклеотидной последовательности соответствующей ДНК и, как правило, объектом синтеза являются гены, кодирующие относительно небольшие белки (например, соматостатин – 14 аминокислот).
2. Выделение полноразмерных генов из клонированных последовательностей ДНК с помощью зондов, созданных на основе химического синтеза (50 – 180 оснований).
Представляет собой выделение и амплификацию индивидуальных последовательностей генов путём репликации в бактериальной системе. Одним из методов является выделение кДНК, т.е. структурной части гена, который предшествует получение индивидуальной иРНК:
иРНК составляет 1 – 5% всей клеточной РНК (80% - рибосомная и 10 – 15% - транспортная) и на 3′ - конце имеет polyA – фрагмент, длина которого достаточна для очистки иРНК с помощью аффинной хроматографии на олиго- (dT)-целлюлозе. Получаемая гетерогенная популяция молекул иРНК кодирует белки, синтезируемые в данный момент клеткой. Затем обычно иРНК фракционируют по размеру при помощи центрифугирования в градиенте плотности сахарозы и изучают на наличие интересующих последовательностей при помощи трансляции in vitro (в лизатах, полученных из зародышей пшеницы или ретикулоцитов кролика, и содержащих все компоненты, необходимые для синтеза белков). Продукты трансляции анализируют при помощи электрофореза на SDS – ПААГ и так как при трансляции in vitro использовались радиоактивно меченные аминокислоты – это позволяет идентифицировать их авторадиографией.
Обнаруженный нужный транскрипт иРНК копируют в ДНК с помощью обратной транскриптазы, образовавшуюся ДНК (кДНК) включают в плазмиды и вводят в бактерии путём трансформации. Клоны бактерий, несущие участок чужеродной ДНК, отбирают из многочисленного окружения нетрансформированных клеток путём гибридизации in situ с радиоактивно мечеными пробами (примерно 15 нуклеотидов).
Представляет собой совокупность всех нуклеотидных последовательностей хромосомной ДНК организма в виде фрагментов ДНК, клонированных в векторах. Если фрагменты физически картированы, т.е. установлено их расположение на хромосомах, то такую библиотеку называют геномной энциклопедией. Банки служат источником материала для изучения строения и регуляции индивидуальных генов, а также для исследования структуры и функции белков. С их помощью также можно решать проблему генофонда исчезающих видов.
Для создания банка генов осуществляют тотальное клонирование, используя для встраивания в вектор смесь фрагментов ДНК, полученную дроблением всей клеточной ДНК. Далее тем или иным способом отыскивают в банке клон с нужным геном.
Банком кДНК называют коллекцию кДНК-копий иРНК клеток конкретной ткани на определённой стадии развития организма. Типичная клетка содержит от 10 до 30 тысяч разных иРНК со средним размером 1,5 – 2 тысячи нуклеотидов, треть из которых представлена низким числом копий (менее 10 – 15 на клетку). Именно эти редкие иРНК определяют, какое число клонов кДНК необходимо отобрать, чтобы банк был представительным. Для вероятности 99% это число близко к 200 000.
Изучение кДНК может дать информацию о структурной части гена, кодирующего какой-либо белок. В это же время полная структура гена, включающая регуляторные последовательности, может быть опознана путём изучения геномных клонов. Сравнительный анализ кДНК-овых и геномных клонов позволит определить наличие интронов или иных последовательностей, отсутствующих в иРНК.
Для клонирования кДНК обычно используют плазмиды, способные включать в свой состав относительно небольшие участки чужеродной ДНК. Клонирование же протяжённых фрагментов геномной ДНК часто осуществляется при помощи бактериофага 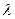 . Часть его ДНК не существенна для жизнедеятельности и репродукции фага и может быть заменена, в связи с чем такие конструкции носят название векторов замещения.
. Часть его ДНК не существенна для жизнедеятельности и репродукции фага и может быть заменена, в связи с чем такие конструкции носят название векторов замещения.
Высокомолекулярную ДНК частично гидролизуют рестриктазой до фрагментов примерно 20 000 полинуклеотидов. В свою очередь ДНК фага разрезают фрагментом рестрикции на 3 части и разделяют электрофорезом в геле. Среднюю, несущественную, часть удаляют, а «плечи» вектора соединяют с геномной ДНК. Затем проводят упаковку фага, инфицируют E.Coli для амплификации ДНК и отбирают соответствующие клоны.
При объединении фрагментов ДНК (например, для клонирования в векторе) возникает проблема совместимости концов объединяемых фрагментов.
Т.к. клонирование необходимо проводить ориентировано – у вектора и у фрагмента геномной ДНК – (у лигируемых молекул ДНК) проводят ферментативную модификацию концов. Сначала для предотвращения циркуляризации обрабатывают щелочной фосфатазой E.Coli, после чего концы таких молекул не замыкаются друг с другом лигазой из-за отсутствия у них фосфатных групп.
Проблему же несовместимости концов фрагмента и вектора решают либо путём достраивания липких концов, либо с использованием линкеров – однонитевых самокомплементарных олигонуклеотидов, которые образуют дуплексы. Имеющие ровные концы и содержащие сайты рестрикции.
Линкеры с определённым сайтом рестрикции используют для присоединения фрагментов ДНК, имеющих ровные концы, к ступенчатым концам вектора, образованным одноимённой рестриктазой. Для этого линкеры сначала «пришивают» с двух сторон к фрагментам кДНК с помощью ДНК-лигазы фага Т4, а затем обрабатывают соответствующей рестриктазой. Далее фрагменты и векторы объединяют через образовавшиеся липкие концы.
Например:
|
5′ CCGAATTCGG…NNNNN……..NNNNN…CCGAATTCGG 3′
 3′ GGCTTAAGCC…NNNN……….NNNN..….GGCTTAAGCC 5′
3′ GGCTTAAGCC…NNNN……….NNNN..….GGCTTAAGCC 5′
 | |||||
| |||||
| |||||
5′ AATTCGG…NNNN…… NNNNN...CCG 3′
3′ GCC…NNN……… NNNN….GGCTTAA 5′
Основная проблема, возникающая при создании банка генов – необходимость обеспечения его полноты. Она определяется вероятностью встречаемости (р) искомого гена в коллекции, состоящей из n клонов:
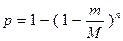 , где
, где
М – молекулярная масса генома;
m – средняя молекулярная масса клонируемых фрагментов ДНК.
К банкам генов условно относят такую коллекцию клонов, в которой индивидуальные гены представлены с вероятностью не менее 0,95.
Число клонов, содержащих рекДНК, которые необходимо отобрать для обеспечения заданной вероятности:
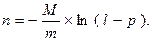
Например, для человека со средним размером гаплоидного генома 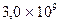 и пшеницы со средним размером гаплоидного генома
и пшеницы со средним размером гаплоидного генома 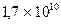 количество клонов для векторов λ при р = 0,99 равно соответственно 690 000 и 3 900 000 (средний размер вставок – 20 000 полинуклеотидов).
количество клонов для векторов λ при р = 0,99 равно соответственно 690 000 и 3 900 000 (средний размер вставок – 20 000 полинуклеотидов).
Существует несколько методов скрининга, т.е. поиска в геномной библиотеке или в банке кДНК тех клонов (бактериальных или фаговых), которые содержат рекДНК с заданным геном. Из них основные – гибридизационный, иммунологический, генетический и рекомбинационный. Общий метод скрининга – гибридизация рекДНК с зондами, т.е. с нуклеотидными последовательностями (олигонуклеотидами, однонитевыми кДНК или иРНК), которые комплементарны отыскиваемому гену. На практике для этого используют метод гибридизации колоний (перепечатка, лизис, авторадиография). Если клонируемые гены экспрессируются (некоторые банки кДНК), то для скрининга применяют иммунологические и другие методы, регистрирующие присутствие специфического белка.








