 2014-02-10
2014-02-10 7424
7424Если освещать растение ярким светом, а затем быстро поместить в темноту, то в первые минуты наблюдается усиление поглощения листьями кислорода и выделение углекислоты. Это светоактивируемое выделение СО2 и поглощение О2 листьями получило название фотодыхания. Оно характерно для растений С3- типа фиксации углекислоты и внешне не обнаруживается у растений с С4-типом.
Если учитывать фотодыхание, то на свету в листьях С3-растений одновременно идут три различных процесса: фотосинтез; митохондриальное (темновое) дыхание; фотодыхание. В связи с этим для получения истинных значений фотосинтеза необходимо принимать во внимание не только темновое дыхание, но и фотодыхание.
Фотодыхание обнаружено и у древесных растений. Этот процесс наблюдался у разных видов сосны, ели, лиственницы, пихты, различных видов и клонов тополя, тюльпанного дерева и других представителей древесного мира.
Фотодыхание осуществляется в особых микротельцах - пероксисомах, имеющих плотную гранулярную строму, окруженную простой мембраной. В них сосредоточены гликолатоксидаза, каталаза, глютамат- и глиоксилаттрансаминаза и другие ферменты, катализирующие превращения веществ в процессе фотодыхания. В ходе этого процесса развертывается довольно сложная картина различных превращений гликолевой кислоты, одного из промежуточных продуктов фотосинтеза.
В присутствии кислорода на свету РДФ-карбоксилаза присоединяет к РДФ не СО2, а О2. Присоединив кислород, РДФ распадается на одну молекулу ФГК и одну молекулу фосфогликолевой кис лоты, которая, отщепляя остаток фосфорной кислоты, превращается в гликолевую и из хлоропластов переходит в пероксисомы. Здесь гликолевая кислота окисляется в глиоксилевую с образованием пероксида. Образующаяся перекись водорода распадается на воду и свободный кислород. Глиоксилевая кислота, реагируя с глутаминовой кислотой, трансаминируется в глицин, который покидает пероксисомы и в митохондриях превращается в другую аминокислоту - серин: из двух молекул глицина образуется одна молекула серина и выделяется одна молекула СО2. Вследствие этого часть зафиксированного в фотосинтезе углерода теряется растением.
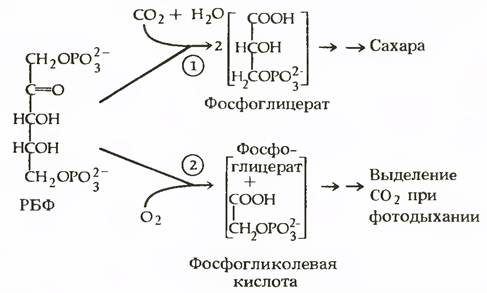
Фотодыхание теснейшим образом связано не только с отдельными звеньями фотосинтеза, но и с темновым дыханием.
До сих пор физиологическая роль фотодыхания полностью неясна. Предполагается, что в пероксисомах клеток листа С3-растений наличие гликолатного метаболизма способствует активированию диффузии бикарбоната в хлоропласт и повышению концентрации СО2, идущей на реакции карбоксилирования по циклу Кальвина. В этой связи фотодыхание следует признать позитивным явлением, свидетельствующим о тесной связи фотосинтеза и окислительных процессов на свету. Часть выделяющейся при фотодыхании углекислоты вновь поступает в хлоропласты, где и фиксируется вновь. Фотодыхание - один из путей синтеза азотистых соединений (глицина).

Фотодыхание выполняет и определенную регуляторную функцию: окисляя часть ассимилятов при отсутствии запроса на них растения, восстанавливают донорно-акцепторные связи. У древесных растений интенсивность фотодыхания ниже интенсивности темнового дыхания. Кроме того, между величиной фотосинтеза и фотодыхания в период активного роста дерева прямой связи не наблюдается, что свидетельствует о возможности предотвращения сильного затоваривания хлоропластов. Этому у древесных растений способствует наличие такого сильного аттрагирующего центра, притягивающего ассимиляты, каким является камбиальная зона дерева. Потребность в ассимилятах на рост дерева по диаметру в значительной степени исключает затоваривание хлоропластов и тем самым исключает необходимость сброса чрезмерного избытка энергии через фотодыхание (Н.Е. Судачкова, Г.И. Гирс, С.Г. Прокушкин и др.). Вместе с тем окисление фотосинтетических продуктов, в данном случае гликолата, можно рассматривать в качестве потерь энергии и метаболитов, только что синтезированных в процессе фотосинтеза. Известно, что у С3-растений фотодыхание может составлять в отдельных случаях 40 % и даже более от чистого поглощения СО2.
Удалось показать видовую специфику фотодыхания у древесных растений. Доказано также, что интенсивность фотодыхания тесно связана с интенсивностью фотосинтеза: с повышением последней увеличивается первая.
Так, по данным Г.М. Голомазовой и др., при насыщающих фотосинтез освещенностях интенсивность фотодыхания лиственницы сибирской составляла 0,60 мг СО2 на 1 г хвои в час, сосны сибирской - 0,42, сосны обыкновенной - 0,40, ели обыкновенной - 0,21. Видно, что наибольшей интенсивностью фотодыхания обладает хвоя лиственницы. При этом необходимо указать, что доля фотодыхания от фотосинтеза у исследованных древесных растений различна: фотосинтез лиственницы и сосны сибирской примерно в 1 раз превышал фотодыхание, сосны обыкновенной - в 5 и ели - в 3 раза.
Интенсивность фотодыхания в значительной степени зависит не только от силы света, но и от температуры воздуха, содержания кислорода и углекислоты, водообеспеченности клеток мезофилла листа и ряда других внешних и внутренних факторов. В темноте данный физиологический процесс полностью отсутствует.








