 2015-02-14
2015-02-14 1810
1810Д’Адамо П., Уитни К. в [2090] утверждают, что «первые носители второй группы крови появились среди кавказских народностей в период между 25 тыс. и 15 тыс. до н.э. в районе западной Азии или Ближнего Востока».
В этой связи интересно отметить, что ни одного кавказского народа, сложенного неоантропами и относящегося к указанной древности, науке не известно. К 10-му тыс. до н.э. население Леванта (Южного Кавказа и прилегающих областей) вообще деградировало практически до нулевой отметки [1737].
Однако на Кавказе в массовом порядке и с самой глубокой древности известны пещерные стоянки неандерталоидов, и вообще регион известен, как один из центров неандертальского заселения (стоит отметить, что и по сей день автохтонные племена Кавказа имею рост менее 164 см – обычный для неандертальского человека) (см. п. 10.2.3. гл. III). Известные на Кавказе стоянки неандертальцев, охотников и скотоводов, относящиеся и к указанной выше дате.
Баски, относящиеся к кавказоидной расе, но имеющие преимущественно первую группу крови (0), возможно, и перешли намеренно в режим изоляции – как европеоидный народ, не желавший скрещиваться с неандертальцами Кавказа.
| % | Страна: народы |
| Индейцы южной Канады и Монтаны: племена блад и черноногих [2136] | |
| Дункеры (США, эмигранты из Германии начала 18-го века) [2136] | |
| 45,2 | Венгрия: венгры 45,2% [2138] |
| 44,7 | Великобритания 42%: англичане 44,7% [2138] |
| Франция | |
| Дания | |
| Финляндия | |
| Швеция | |
| 42,1 | Нидерланды: голландцы 42,1% [2138] |
| Канада | |
| 40,3 | Австралия 38%: австралийцы 40,3% [2138] |
| США [2138], 40 – 45% [2136] | |
| Бельгия | |
| Польша | |
| 36,7 | Япония: японцы 36,7% [2138] |
| Россия [2135]: русские 35,8% [2138] | |
| 34,5 | Южная Корея |
| Арабы [2138] | |
| Эскимосы [2136] | |
| 22,6 | Гонконг, Китай 26%: китайцы 22,6% [2138] |
| Бенгалия: индийцы 24,5% [2138] | |
| Эскимосы Гренландии [2136] | |
| Индейцы Центральной и Южной Америки [2094] |
Таблица 4.8.1.2.1. Распределение второй группы крови (А) (среднее в популяции,%).
Регион с большим содержанием второй группы крови (А) – Великобритания – Франция – Дания – Нидерланды – Бельгия – является регионом традиционно древнего заселения неандертальским человеком, ростом менее 164 см (см. п. 4.7.2.0. гл. IV). Некоторые исследователи ошибочно неандертальскую ветвь Южной Европы называют средиземноморской расой, относя её к подрасам европеоидной большой расы. Ошибка заключена и в бравировании еврейских учёных выведением своей наследственности от древнего населения пещеры Шанидар (Левант). Найденные в ней люди были неандертальцами, и по современной теории евреи не могут происходить от неандертальцев.
Заметным маркером второй группы крови (А) может служить Венгрия с её 45,2%, поскольку венгры относятся не к европеоидному населению, а к уральскому (тюркскому). В этногенезе финнов и шведов также заметную роль сыграли уральские (тюркские) племена.
Другие исследователи относят появление второй группы крови (А) ко времени неолита (8 – 3 тыс. до н.э.) и связывают изменение состава крови с переходом человека к земледельческому образу жизни и произошедшими в питании людей изменениями, повлёкшими за собой мутации пищеварительной и иммунной систем. Однако против такого предположения свидетельствуют своей кровью аборигены Австралии, имеющие 40,3% крови группы А, но до сих пор не перешедшие к земледельческому образу жизни. И эскимосы с 30% никогда ничего не выращивали в зонах многолетней мерзлоты.
Распространение земледелия имело общий вектор, направленный из Русской равнины в сторону Передней Азии, Кавказа и Востока вообще. Поэтому, если связывать возникновение второй группы крови (А) с земледелием, то вторая группа крови (А) должна иметь максимум распространения среди европейских народов среднего пояса, привязанных к наиболее плодородным землям планеты – средняя часть Русской платформы.
Однако данные, представленные в таблице 4.8.1.2.1., дают нам другие сведения. Максимум носителей второй группы крови (А) в 44% относится к западным и северо-западным областям Европы и снижается до 22,6% в Китае, 26% в Гонконге, и даже до 20% в Бенгалии (в бассейне нижнего течения Ганга и дельты Ганга и Брахмапутры в восточной части Индии). При этом Дания (44%) отнюдь не родина земледелия, а экономики Китая и Индии в большей степени основаны на земледелии.
Другие исследователи пытаются искать связь между преобладанием второй группы крови (А) и становлением городов. По этим данным, во многих областях, имеющих долгую историю градостроительства, там, где часто случались вспышки чумы, холеры и оспы, отмечается преобладание второй группы (А) над первой (0). Однако даже самих исследователей удивляет тот факт, что первая группа крови (0) вообще устояла в таких условиях, и более того, до сих пор остается самой распространённой на планете.
Между тем, вторая группа крови (А2) также широко распространена в Исландии и Скандинавии, преимущественно среди саамов, пришедших на Кольский полуостров и север России в 1-ом тыс. до н.э. Общая их численность в начале 20-го века составляла около 33000 человек, из которых 68,9% жили в Норвегии, 21% – в Швеции [51]. При этом саамы этнически достаточно изолированы. Основными занятиями саамов являлись оленеводство, рыболовство, морская и сухопутная охота. Традиционная пища cаамов зимой состояла из оленьего мяса, летом – из рыбы.
Следовательно, имея высокую степень распространения гена второй группы крови (А), саамы и австралийцы, абсолютно неземледельческие народы, опровергают теорию зависимости выработки второй группы крови (А) от перехода человека к земледелию.
Язык саамов принадлежит к финно-угорской группе языков уральской семьи, родиной которой являются западные районы Юго-Восточной Азии, а этногенез носителей связан с уральской расой (ветвь монголоидной расы). Отсюда можем предположить, что поставщиком гена второй группы (А) является монголоидная составляющая (плечо «сино-» в синокавказской общности) в крови саамов. И именно с ней связана высокая процентная доля носителей второй группы крови (А) среди народов Кольского полуострова. При этом большая часть монголоидов уральских и кавказских очагов являются народами неандертальского происхождения (рост ниже 164 см).
Поэтому ген А2 (разновидность гена А) является «визитной карточкой» кавказских народов. Вторая группа крови (А) в значительной степени представлена в Средиземноморском бассейне и, главным образом, на Корсике, в Сардинии, Испании, Турции, на Балканах. Обратим наше внимание на то, что этногенез перечисленных территорий связан с многовековым присутствием тюрко-язычного населения – пришлого в виде различных каганатов и автохтонного неандертальского.
Первые четыре указанных региона связаны с турецким этносом, который непосредственно связан с тюрками – Брокгауз и Ефрон в статье «Турки» прямо отсылают читателя к статье «Тюрки» [47], а самоназвание турков – тюрк [35]. Этнически турки сложились из двух основных компонентов: тюркских кочевых скотоводческих племён (главным образом евреев-огузов, народа тюркского племени), переселившихся в Малую Азию из Средней Азии и Ирана в 11 – 13 вв., во время монгольских и сельджукских завоеваний, и местного малоазийского населения. Часть тюркских племён проникла в Малую Азию с Балкан (узы и печенеги). Смешиваясь с местным населением (греками, армянами, грузинами и др.), тюрки ассимилировали его [2123]. Греки к началу нашей эры уже в большинстве своём были представлены семито-кавказоидами (см. п. 7.1.2.2. гл. IV), носителями синокавказских языков. Население Балкан – узы, печенеги, болгары, авары и т.д. – также относятся к тюркам (с общественным укладом в виде каганатов, то есть иудеи).
Среди азиатов только в Японии и в субконтинентальной Индии существуют также области с большим числом носителей второй группы крови (А). При этом японцы сформировались как группа племён в середине 1-го тыс. до н.э. в результате переселения группы носителей энеолитической Яёи культуры с юга Корейского полуострова на Японские острова, населённые до этого айнскими и частично малайско-полинезийскими племенами [603]. Айны (см. рис. 4.8.1.2.1.) по языку и физическому типу обнаруживают сходство с населением Юго-Восточной Азии, оттуда их предки в раннем неолите мигрировали в Японию. Обнаруживая сходство с выше обозначенными саамами: традиционный уклад жизни айнов, как и саамов, основан на осёдлом рыболовецком и охотничьем хозяйстве. Айны заселяли не только всю Японию, но и низовья Амура, Камчатку, Сахалин и Курильские острова. В расовом отношении айнов относят к австралоидам, а также к тунгусам, алтайцам и другим жителям Урала и Сибири [961].

Рис. 4.8.1.2.1. Корреляция дояпонских айнов и австралоидов с семито-кавказоидами:
1 – изображение айна с острова Эзо (худ. Скотников Е.О., ок. 1804 г.); 2 – айн (1094 г.); 3 – еврей (К. Маркс, 19 в.); 4 – айн (австралоид); 5 – айн (1904 г.); 6 – еврей (раввин, 20 в.); 7 – кавказец (20 в.).
В Европе обладатели второй группы крови (А) чаще всего встречаются среди западных европейцев, куда из Африки и с Балкан постоянно проникали тюркско-арабские элементы. По мере продвижения на восток Европы – в пределы расселения русского населения (и других чисто славянских и чисто европеоидных племён) – степень распространения второй группы крови (А) уменьшается.
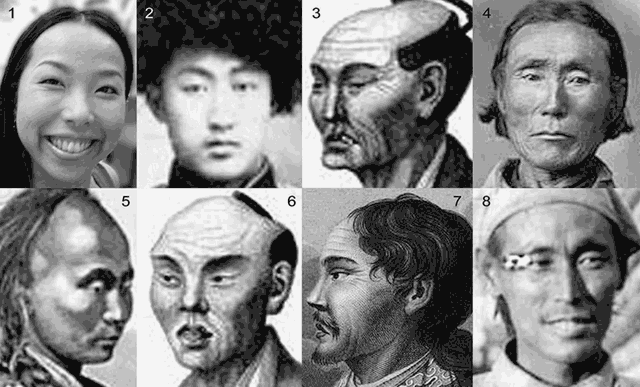
Рис. 4.8.1.2.2. Представители народов второй группы крови (А):
1 – современная айнка (Канако Узава, Япония, 2007); 2 – кореец (в казацкой форме, г. Николайск-Уссурийский, нач. 20-го в.); 3, 6 – изображение японцев (худ. Масловский П.И., нач. 20-го в.); 4 – эвенк (18 в.); 5 – гольд (нанаец, 1985 г.); 7 – татарин (худ. Скотников Е.О., нач. 20-го в.); 8 – енисеец (арины/арий).
Для 10% от общего числа евреев, принадлежащих к арменоидному типу балкано-кавказской расы, самой характерной является также вторая группа крови (А) [2192]. А, ввиду того, что оставшиеся 90% евреев принадлежат к тюркам (хазарский, тюркский, болгарский, огузский, аварский и др. каганаты; которые не входили в число земледельческих народов, а занимались преимущественно кочевым образом жизни, см. гл. XI), можем и здесь предположить, что арменоидные евреи получили вторую группу крови (А) от своих азиатских предков.
«В глубокой древности австралоидный, антропологический тип был широко распространён в Южной и Юго-Восточной Азии. Наибольшее сходство с австралийской расой имеют ведды Цейлона (цейлоно-зондская раса) и южно-индийская (дравидийская) раса» [35, ст. Австралоидная раса]. Вектор переселения австралийцев известен – из Индостана в Австралию, около 30-го тыс. до н.э. Поэтому высокие проценты второй группы крови (А) среди венгров (угорские племена, родственные манси и хантам), а также среди австралийских аборигенов (австралоидная раса), японцев, финнов и т.д. говорят о том, что ген второй группы кровы (А), скорее всего, связан с австралоидной составляющей в расогенезе, в том числе и в образовании тюрок.
К тюркским народам относятся народы Азии и Восточной Европы: якуты, эвенки, татары, алтайцы, сойоты, долганы, таранчи, хами, иегуры, киргизы, узбеки, чуваши, тептяри, башкиры, туркмены, сарты, турки и др. Тюрки имеют кавказский или близкий к нему тип (эти и дают высокие показатели крови второй группы (А) среди народов Кавказа), другие (восточные тюрки, живущие близ Монголии и внутри неё) имеют монгольский или близкий к нему тип [47].
К 1970 году в СССР тюрки-уральцы достигли численности 62.900.000 человек, то есть более 25% всего населения СССР, остальные 75% приходилось на русских – 129.015.100, украинцев – 40.753.100, белорусов – 9.051.800.
Поэтому средний показатель для России 36% второй группы крови (А) завышен, по крайней мере, на четверть за счёт тюркского населения, то есть с учётом этой поправки среди русского населения России будет 43% носителей первой группы крови (0) и 25% носителей второй группы крови (А). Что по показателю первой группы крови (0) ставит Россию в обще-западноевропейский ряд.
Таким образом, можем сделать следующие выводы:
- Обладание второй группой крови (А) находится в зависимости от присутствия в этногенезе рассматриваемого народа тюркско-уральской составляющей – совпадает с ареалом расселения тюркско-уральских народов.
- Вторая группа крови (А) распространена преимущественно в зоне смешения местных европеоидных народов и пришлых тюркско-уральских.
- Местом генерации второй группы крови (А) является регион начального распространения представителей уральской расы – от западной (Тибет) до северо-восточной оконечности (Корея) Юго-Восточной Азии.
- Низкие проценты носителей второй группы крови (А) в самом Тибетско-Корейском регионе говорит о том, что эта группа крови связана именно с местом контакта тюрок и европеоидов, а не с теми регионами планеты, в которых тюрки являются автохтонным или преимущественным населением.
- Учитывая то, что монголоидные американские индейцы и австралоидные австралийские аборигены обладают первой группой крови (0), можем заключить, что принадлежность тюрок к монголоидной или айнов к австралоидной расам не является необходимым условием для возникновения второй группы крови (А).
- Монголоидная раса не является генератором второй группы крови (А), но служит её поставщиком в районы расселения европеоидов.
- Учитывая то, что американские индейцы прекратили общаться с Евразией около 10 тыс. до н.э., обнаруживаем, что индейцы вышли из Евразии без гена второй группы крови (А), то есть, возможно, до 10-го тыс. до н.э. монголоиды не смешались с носителями второй группы крови (А).
- Уже к 30 – 20-ому тыс. до н.э. вторая группа крови (А) существовала в районе распространения веддидов и дравидов, поскольку этими племенами в это время была заселена Австралия, где сохранился высокий процент второй группы крови (А).








