 2015-03-22
2015-03-22 1091
1091Гл а в а 2
ЛАНДШАФТЫ ЛЕСНОЙ ЗОНЫ РУССКОЙ РАВНИНЫ
И ФАКТОРАЛЬНО-ЭКОЛОГИЧЕСКИЕ РЯДЫ
БИОГЕОЦЕНОЗОВ
Дифференциация лесных биомов: общая
Схема
Итак, в предыдущей главе мы попытались представить, как появился субстрат, своего рода арена для развития современных ландшафтов Русской равнины. Освободившаяся ото льда пример-но 13—15 тыс. лет назад территория являла взору древнего челове-ка весьма суровый пейзаж. Высокие моренные водоразделы еще сохраняли реликтовую мерзлотность, небольшие островки леса из березы, сосны и ели разделялись широкими пространствами тундростепи.
Ранние этапы послеледниковья отмечены преобладанием бе-резовых лесов, конкуренцию которым составляла селившаяся на песчаных грунтах сосна: березняки занимали моренные возвышен-ности, а в долинах рек и на обрамляющих их зандровых равнинах господствовали сосняки. Так впервые заявила о себе биота ланд-шафта — его живая «пленка», тонкий слой, охватывающий всего лишь несколько первых десятков метров — от ближайшего к по-верхности горизонта подземных вод до верхушек крон деревьев первого яруса древостоя.
Становление биоты ландшафта сопровождалось непрерывны-ми волнообразными изменениями климата. Потепления, отогре-вавшие поверхность и приводившие к вытаиванию линз погре-бенного льда, сменялись похолоданиями, приносившими новые волны холода, вслед за которыми восстанавливался приледни-ковый растительный комплекс, задерживалось развитие лесных сообществ. И все же вторжение последних на север было предоп-ределено общей направленностью развития биосферы. Основные виды — строители биоценозов — приняли старт примерно около 11 тыс. лет назад и начали проникать на север, осваивая разные формы рельефа. Ель со скоростью 350 — 500 м в год продвигалась по глинистым моренным холмам, еще сохранявшим остаточную «вечную» мерзлоту. Сосна двигалась медленнее — по террасам рек, супесчаным водно-ледниковым равнинам, преодолевая око-ло 200 — 300 м в год. Береза составила конкуренцию ели и дубу,
всюду, где позволяли условия, примешиваясь к хвойным древо-стоям.
При этом оказалась, что виды — «строители» биоценозов (на-зовем их эдификаторами, как это принято в экологии) — прояв-ляют совершенно четкую «привязанность» к характеру грунта, достигая господства и образуя монокультурные, т.е. состоящие из одной породы леса в определенных условиях: сосна — на песках и супесях, ель — на глинах и суглинках. Таким образом, механиче-ский состав четвертичных отложений выступил в роли главного об-стоятельства, обусловившего видовой состав коренных биогеоцено-зов Русской равнины. И это неудивительно, ведь механический со-став грунтов определяет целый комплекс параметров, характери-зующих каждое отдельное местообитание, а именно:
• богатство субстрата, или его трофность (гр. trophe — «пита ние») — наличие в грунтах макроэлементов и микроэлементов, необходимых для питания и роста растений;
• водно-воздушный режим формирующихся на данном субстрате почв, т. е. их проницаемость для воды и воздуха, способность «ды шать», пропускать и задерживать влагу;
• температурный режим — глубину прогревания-промерзания и, что немаловажно, скорость, с которой грунты отогреваются весной;
• проточность или, напротив, застойность атмосферных осад ков и талых весенних вод.
По своему воздействию на формирование ландшафтов все раз-нообразие четвертичных отложений может быть в первом прибли-жении сведено в две большие группы:
1) моренные суглинистые грунты, на которых развиваются тяжелые по механическому составу почвы, а доминантой (преоб ладающей породой) первого яруса древостоя выступает ель;
2) водно-ледниковые супесчаные и песчаные наносы с легки ми почвами и сосной в качестве доминанты первого яруса.
Есть еще сложносоставные грунты, состоящие из двух слоев — одного глинистого и одного супесчаного. Почвы, формирующи-еся в таких условиях, называются почвами на двучленах. Ландшаф-ты на двучленах занимают как бы промежуточное положение. Су-песи, залегающие на суглинках, дают примесь сосны к еловому древостою, примесь тем более заметную, чем больше мощность покровного чехла, которая обычно колеблется от 0,5 до 3 м. По-кровные суглинки близки по своему составу к моренам и, следо-вательно, обеспечивают преимущество ели в первом ярусе.
Для широколиственных пород, входящих в состав первого яру-са в подзоне хвойно-широколиственных лесов, оказались весьма значимыми два показателя местообитания: трофность грунтов, во-первых, и средние температуры зимнего месяца, во-вторых. По-этому широколиственные породы (дуб, вяз) распространялись по
богатым моренно-карбонатным грунтам возвышенностей и пой-мам рек, причем их продвижение на север началось значительно позднее, на пороге VI тысячелетия до н.э. во время очередного значительного потепления — климатического оптимума (благо-приятный для широколиственных пород теплый и влажный пери-од), который продлился до III тысячелетия до н.э. Новая волна холода, прокатившаяся 5 тыс. лет назад по всему северному полу-шарию, выбила из древостоев широколиственные виды, прежде всего вяз.
Окончательное установление господства темнохвойной тайги связывают с похолоданием и увлажнением климата около 3,2 тыс. лет назад. Многоярусные дубравы атлантического времени (суще-ствовавшие в период климатического оптимума значительно се-вернее современной Москвы — на склонах Клинско-Дмитровской возвышенности и в бассейне озера Плешеево) постепенно пере-рождаются в более привычные для нас на этих широтах елово-широколиственные леса.
Ныне широколиственные древесные породы практически ниг-де в природе не пересекают границу, совпадающую с январской зимней изотермой -10 °С. Севернее этой изотермы дубы и липы, вязы и клены в составе древостоев первого яруса встречаются только в культурном ландшафте. Этим обстоятельством и объяс-няется наличие природного рубежа, делящего огромный и не-когда сплошной биом — леса Восточно-Европейской равнины на две крупные природные ландшафтные зоны: зону тайги (тем-нохвойных лесов) и зону хвойно-широколиственных лесов (после-днюю иногда еще называют подтаежной). Ареал распростране-ния собственно широколиственных лесов ограничивается обла-стью теплых влажных зим морского климата и до центральных и северных регионов России не доходит, вклиниваясь на террито-рии стран Балтии.
Таким образом, центр Русской равнины занят хвойно-широ-колиственными лесами, в которых дуб и липа примешиваются к основному древостою из ели и сосны. Север — царство тайги, протянувшейся широкой полосой от ярославских, костромских и вятских лесов до тундр Кольского полуострова. Тайгу еще называ-ют бореальной лесной зоной (от лат. borealis — «северный») Лан-дшафтоведы и геоботаники различают в пределах зоны тайги три подзоны, различающиеся характером растительных сообществ: северная тайга, средняя тайга и южная тайга.
Сразу отметим: путешествуя с юга на север (скажем, из Моск-вы в Вятку), мы нигде на найдем четкой границы между хвойно-широколиственными и таежными лесами, во всяком случае в виде линии, севернее которой нельзя встретить представителей широ-колиственных пород. Напротив, дуб, липу или клен можно обна-ружить в подросте и втором ярусе естественных древостоев и го-42
раздо севернее пределов распространения хвойно-широколиствен-ных лесов, например на территории Ярославской и Вологодской областей. Лесникам Заволжья хорошо известно, что следующие подряд одна за другой на протяжении пяти-шести лет теплые зимы приводят к появлению в тайге под покровом ели заростков дуба. Следовательно, при благоприятных условиях широколиственные породы как бы диффузно проникают на территорию более север-ной природной зоны, распространяясь по всей «приграничной» полосе.
Северная граница тайги также представляет собой весьма плав-ный переход к тундре, и формальная линия границы лесов может быть проведена лишь условно. Изреживание лесного покрова про-исходит постепенно — от более или менее сомкнутых северо-та-ежных лесов до редколесий, фактически представляющих собой отдельные деревья и группы деревьев в тундре. Дальше всего сом-кнутые леса заходят в тундру по долинам рек, отдельные неболь-шие лесные массивы и островки встречаются также по наиболее прогреваемым или защищенным от сильных северных ветров фор-мам рельефа. Болота, занимающие обширные водораздельные пространства и речные террасы вблизи северного предела распро-странения лесной растительности, фактически образуют единое нелесное пространство с зональными (климатически обусловлен-ными) тундрами.
В целом несмотря на общепринятое мнение о том, что тайга Европейской России характеризуется четко выраженной зональ-ностью, сколько-нибудь четкие и однозначно признаваемые гра-ницы между притундровой, северной, средней и южной тайгой отсутствуют. Различия в пределах, например, средней тайги, свя-занные с особенностями геологического строения местности, макрорельефом и составом четвертичных отложений, существен-но превышают различия между аналогичными по этим показате-лям участками средней и северной или средней и южной тайги. Кроме того, имеющиеся данные о распространении древесных пород в разные периоды голоцена по территории Европейской России показывают, что относительно четкое разделение расти-тельности на природные зоны по составу лесов произошло со-всем недавно и, возможно, является в значительной степени след-ствием зональности хозяйственной деятельности. Таким образом, можно утверждать, что лесная зона России представляет собой сложный природно-антропогенный комплекс, в формировании и развитии которого уже на протяжении многих столетий хозяй-ственная деятельность человека играет решающую роль.
Примеры ландшафтных описаний и полевые наблюдения, по-ложенные в основу этой книги, в основном охватывают южную тайгу и хвойно-широколиственные леса, во-первых, потому, что именно здесь когда-то зарождалась Северо-Восточная Русь, во-вторых, просто потому, что автор лучше знаком с ландшафтами этих подзон.
Но, довольно теории — попробуем совершить путешествие по основным ландшафтам южно-таежной зоны. Впрочем, в данном случае речь пойдет об экспедиции, ибо нам придется преодолеть немалые расстояния в поисках девственного леса, или, точнее, леса, ни разу не вырубавшегося на памяти поколений, такие леса называются условно-коренными.
Начнем с высоких (250 м абсолютной высоты и выше) морен-ных гряд (рис. 2.1),
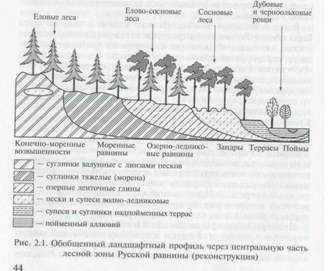
где на одиночных холмах и куполах из валунно-го суглинка воцарилось безмолвие и мрак елового леса. Примесь сосны здесь можно обнаружить лишь над линзами песка среди глин. Как мы сможем убедиться чуть позднее, ельник вовсе не скучен и уж точно не однообразен: характер леса меняется от сумрачного «мертвопокровного», т.е. лишенного травяного или мохового напочвенного покрова, до «веселого» с мягким изум-рудным ковром сфагновых мхов.
Ниже по рельефу (170 — 240 м) располагаются монотонные моренные равнины, где еловая тайга прерывается только отдельны-ми камовыми холмами, поросшими сосной.
Что же представляет собой настоящая темнохвойная тайга? Лесообразующей породой еловых лесов является ель обыкновен-ная (Picea abies) — крупное дерево, достигающее высоты 30 м и более, с прямым стволом и конической кроной. Однако корневая система этого гиганта — приповерхностная «розетка», вследствие чего ель сильно страдает от низовых пожаров и легко подвержена вывалу при сильных ветрах.
«Воспитанная» эволюцией в суровом северном климате ель те-невынослива — густота хвои сохраняется даже и во внутренней части кроны; ель не требовательна к теплу, зимостойка и способ-на селиться на бедных элементами питания грунтах. Пожалуй, «капризна» ель только в одном — не любит сухих местообитаний и предпочитает влажную почву.
Ель — сильный эдификатор, образуя густые сомкнутые насаж-дения, она «выстраивает» биоценоз под себя, поскольку задает световой и ветровой режим в местообитании, изменяет почвен-ные условия и таким образом определяет среду существования для других растений. В ельниках создается сильное затенение и местный микроклимат, характеризующийся ровным ходом суто-чных температур и замедленным теплооборотом между почвой и нижним слоем воздуха.
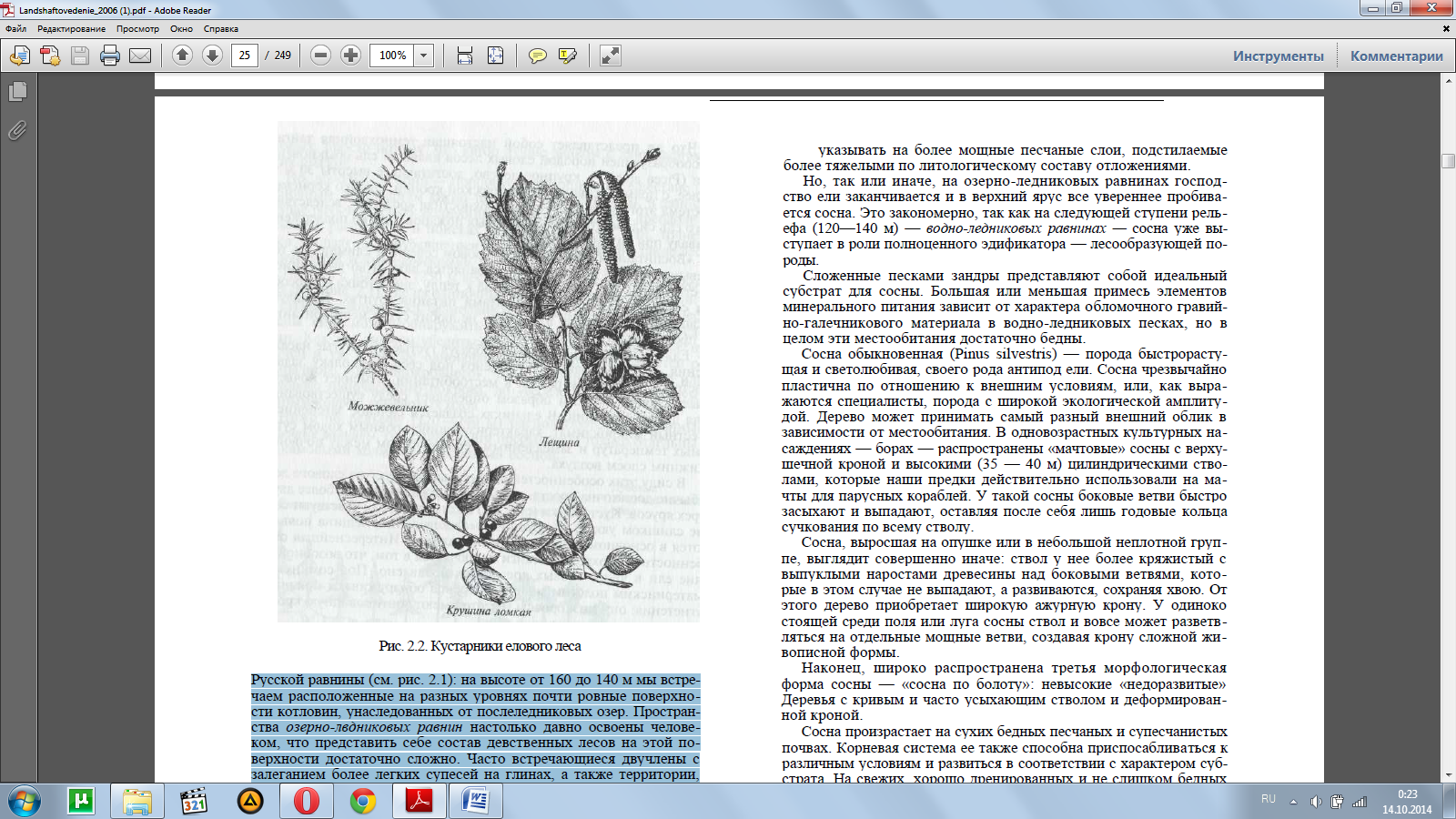
В силу этих особенностей вертикальная структура елового леса обычно достаточна проста, здесь редко можно встретить более двух-трех ярусов. Кустарники (рис. 2.2) под пологом ели чувствуют себя не слишком уютно — крушина, можжевельник, лещина появля-ются в основном в просветах и прогалинах. Интереснейшая осо-бенность темнохвойной тайги заключается в том, что возобновле-ние ели в чисто еловых древостоях подавлено. Под сомкнутым материнским пологом молодые деревца обнаруживают признаки угнетения: они низкорослы и часто имеют зонтиковидную крону, короткую и недолговечную хвою.
Немногие растения способны выдерживать постоянное сильное затенение, поэтому флористическое богатство ельников невели-ко: кислица обыкновенная, майник двулистный, рамишия однобо-кая, одноцветка крупноцветная, плаун булавовидный, ожика воло-систая, вероника лекарственная, голокучник Линнея, грушанка круглолистная, брусника, папоротники. Все эти виды — спутники ели — типичны для южной тайги (их называют бореальными), теневыносливы, требовательны к повышенной влажности воздуха и почв и способны произрастать на небогатых кислых почвах.
Кислыми почвы ельников становятся вследствие многолетнего просачивания атмосферных осадков и талых вод через хвойный °пад. Повышенная влажность, слабая аэрация и кислотность почв ельников способствуют развитию мохового покрова.
Однако пора двигаться дальше, т.е. спускаться ниже по релье-Фу на нашем ландшафтном профиле, проведенном через центр Русской равнины (см. рис. 2.1): на высоте от 160 до 140 м мы встре-чаем расположенные на разных уровнях почти ровные поверхно-сти котловин, унаследованных от послеледниковых озер.
Простран-ства озерно-лвдниковых равнин настолько давно освоены челове-ком, что представить себе состав девственных лесов на этой по-верхности достаточно сложно. Часто встречающиеся двучлены с залеганием более легких супесей на глинах, а также территории, сложенные ленточными озерными глинами, были покрыты сме-шанными елово-сосновыми лесами. Большее участие ели в древо-стое наблюдается на участках с близким от поверхности залегани-ем глин или суглинков, перекрытых менее мощным песчаным плащом. Соответственно господство в древесном ярусе сосны мо-указывать на более мощные песчаные слои, подстилаемые более тяжелыми по литологическому составу отложениями.
Но, так или иначе, на озерно-ледниковых равнинах господ-ство ели заканчивается и в верхний ярус все увереннее пробива-ется сосна. Это закономерно, так как на следующей ступени рель-ефа (120—140 м) — водно-ледниковых равнинах — сосна уже вы-ступает в роли полноценного эдификатора — лесообразующей по-роды.
Сложенные песками зандры представляют собой идеальный субстрат для сосны. Большая или меньшая примесь элементов минерального питания зависит от характера обломочного гравий-но-галечникового материала в водно-ледниковых песках, но в целом эти местообитания достаточно бедны.
Сосна обыкновенная (Pinus silvestris) — порода быстрорасту-щая и светолюбивая, своего рода антипод ели. Сосна чрезвычайно пластична по отношению к внешним условиям, или, как выра-жаются специалисты, порода с широкой экологической амплиту-дой. Дерево может принимать самый разный внешний облик в зависимости от местообитания. В одновозрастных культурных на-саждениях — борах — распространены «мачтовые» сосны с верху-шечной кроной и высокими (35 — 40 м) цилиндрическими ство-лами, которые наши предки действительно использовали на ма-чты для парусных кораблей. У такой сосны боковые ветви быстро засыхают и выпадают, оставляя после себя лишь годовые кольца сучкования по всему стволу.
Сосна, выросшая на опушке или в небольшой неплотной груп-пе, выглядит совершенно иначе: ствол у нее более кряжистый с выпуклыми наростами древесины над боковыми ветвями, кото-рые в этом случае не выпадают, а развиваются, сохраняя хвою. От этого дерево приобретает широкую ажурную крону. У одиноко стоящей среди поля или луга сосны ствол и вовсе может разветв-ляться на отдельные мощные ветви, создавая крону сложной жи-вописной формы.
Наконец, широко распространена третья морфологическая форма сосны — «сосна по болоту»: невысокие «недоразвитые» Деревья с кривым и часто усыхающим стволом и деформирован-ной кроной.
Сосна произрастает на сухих бедных песчаных и супесчанистых почвах. Корневая система ее также способна приспосабливаться к различным условиям и развиться в соответствии с характером суб-страта. На свежих, хорошо дренированных и не слишком бедных почвах у сосны развиваются стрежневой корень и мощная гори-зонтальная корневая система. Такая сосна отличается большой ветроустойчивостью. На сухих, бедных почвах с низким уровнем грунтовых вод и на болотах корневая система сосны поверхност-ная; в этих случаях деревья легко подвергаются ветровалу. Свето-любие и быстрый рост сосны сделали ее породой-пионером, бы-стро заселяющей открытые пространства.
Полевые наблюдения








