 2015-03-20
2015-03-20 651
651Свет является одним из основных климатических факторов, определяющих возможности существования организмов. Только на свету происходит фотосинтез и, следовательно, могут существовать фотосинтезирующие растения. Неподвижным растениям приходится приспосабливаться к определенному уровню освещения. Они могут быть поделены по отношению к свету на три группы: светолюбивые, обитающие в условиях хорошей освещенности, тенелюбивые, обитающие всегда в условиях значительной затененности, и теневыносливые, предпочитающие жить на свету, но могущие существовать и в тени. Подвижные животные, помимо активного выбора местообитания с определенными условиями освещенности, имеют возможность строить свою суточную активность так, чтобы получать определенные количества света. Среди них различают животных дневных, ночных и сумеречных. Будучи активными лишь определенную часть суток, остальное время животные проводят в норах и убежищах, прячутся под пологом листвы, среди лесной подстилки и т. д. Характер активности животных может меняться в зависимости от времени года, метеорологической обстановки и условий освещения. Так, обитающие в полупустынях суслики в жаркие дни бывают активными в утренние и вечерние часы, а околополуденное время проводят в норах; в пасмурные дни они могут появляться на поверхности и днем. Многие птицы пустыни, избегающие в жаркие летние дни появляться на открытых местах, осенью, зимой и весной проявляют максимум активности в дневные часы. Полевки в затененных участках проявляют круглосуточную активность с максимумом в ночные часы, а на освещенных участках активны только ночью.
Растения, выросшие в тени, характеризуются тонкими, нежными листьями с относительно слабо развитой палисадной тканью и хорошо развитой губчатой. У этих растений междоузлия стеблей тонкие, вытянутые, механическая ткань развита слабо. У растений, выросших на свету, листья плотные с хорошо развитой палисадной тканью, стебли толстые, с короткими междоузлиями. Под действием света растения в результате изменения скорости ростовых процессов могут поворачиваться в сторону источника света (положительный фототропизм), в сторону, противоположную источнику света (отрицательный фототропизм), и перпендикулярно направлению световых лучей (трансверсальный фототропизм). Примером последнего является возникновение так называемой листовой мозаики, когда листья располагаются так, чтобы наилучшим образом улавливать солнечные лучи. Наоборот, в тех случаях, когда света много, листья могут свисать с дерева, так что солнечные лучи скользят вдоль их поверхности (например, у австралийского эвкалипта) или обращены ребрами на север и на юг, а плоскостями на запад и восток (у компасных растений аридных районов мира).
Если неподвижные растения реагируют на свет при помощи изменения своего положения по отношению к источнику света — фототропизма, то подвижные животные обладают фототаксисом — они способны двигаться по направлению к свету (положительный фототаксис) или от света (отрицательный фототаксис).
Часть света, падающего на поверхность растений, образующих сообщество, отражается, часть поглощается растениями, а часть проникает внутрь сообщества и там либо поглощается растениями нижних ярусов, либо достигает поверхности почвы. Естественно, что при этом сила света постепенно понижается, а его спектральный состав меняется. Таким образом, растительные сообщества дифференцируют световые условия, характерные для какой-либо местности: неодинаковые условия освещенности создаются в разных сообществах, в разных участках одного и того же сообщества, в одном и том же сообществе в разные сезоны года. Количество света, проникающего под полог деревьев верхнего яруса, связано с относительным минимумом света, т. е. с той величиной его, при которой растение данного вида еще может ассимилировать (рис. 12). Для лиственницы европейской относительный минимум силы света составляет в среднем около 20% от полного дневного освещения, для сосны обыкновенной — около 10, для дуба черешчатого — 3,8, для ели европейской — 3,0, для бука лесного — 1,5, для самшита — 1%. Это значит, что в лиственничном лесу значительно светлее, чем в сосновом, а в сосновом светлее, чем в еловом, дубовом и тем более буковом и самшитовом. Последний является наименее светопроницаемым из лесов, произрастающих на территории СССР. Еще сумрачнее под пологом влажного тропического леса, куда проникает лишь 0,2% полного дневного света. Для лесных травянистых растений освещенность значительную часть дня может быть ниже так называемой компенсационной величины, т. е. такой освещенности, при которой ассимиляция и дыхание уравновешивают друг друга (ниже этой величины растение в принципе существовать не может, так как его дыхание становится более интенсивным, чем ассимиляция). Оказывается, что такое теневое растение, как кислица, может довольствоваться короткими промежутками времени, когда освещенность превышает компенсационную величину благодаря падению на растение солнечного блика (рис. 13).
В лесах, образованных деревьями с опадающей листвой, весной освещенность поверхности почвы велика, и в это время в лесу зацветают некоторые светолюбивые травы. Под пологом хвойного или вечнозеленого леса сезонные изменения освещенности незначительны (рис. 14).
Помимо общего количества света, получаемого организмами, для них имеет большое значение и фотопериодизм, т. е. характер чередования светлых и темных периодов (дня и ночи). Фотопериодизм меняется от приэкваториальных широт к умеренным и холодным поясам. Близ экватора продолжительность дня и ночи мало различается в течение круглого года; в высоких широтах летом день бывает весьма продолжительным, а ночь коротка, зимой — наоборот. Обитатели этих широт — организмы длинного дня, поскольку летом, в период их активности, день значительно превышает ночь; обитатели тропиков — организмы короткого дня. Обитателям разных широт свойственны приспособления к определенному фотопериодизму.
Так, растения длинного дня нуждаются в высокой продолжительности светлого периода, чтобы накопить достаточное количество органического вещества. Если их поместить в условия короткого дня, они будут успешно вегетировать, но не смогут перейти к образованию генеративных органов, так как светлой части суток недостаточно для накопления достаточных запасов органического вещества. Растения короткого дня, перенесенные в условия длинного дня, не смогут перейти от вегетативного развития к генеративному. Это связано с тем, что они на родине в течение более продолжительной ночи успевают освободить листья от накопленных органических веществ и тем самым подготовить ассимиляционный аппарат для успешного фотосинтеза к началу следующего дня, а в низких широтах ночь слишком коротка для этого. У животных умеренного пояса (животных длинного дня) осеннее укорочение дня ведет к прекращению процессов размножения. Весеннее удлинение дня возобновляет способность к размножению. Механизм действия этого удлинения, видимо, таков: удлинение дня действует на гипофиз — придаток головного мозга, тот стимулирует работу эндокринных органов, которые передают поступивший сигнал половым железам. Увеличение длины светового дня путем дополнительного ежесуточного освещения птичников позволяет повысить яйценоскость домашней птицы.
С другой стороны, урожайность яблони — дерева короткого дня — может быть повышена, если в условиях длинного дня закрывать ее крону на несколько часов в сутки деревянным непроницаемым для света шатром. Это одновременно приводит также к повышению морозоустойчивости этого южного дерева, что может служить примером замещения действия факторов, о котором мы говорили ранее.
Одним из наиболее важных климатических факторов, определяющих размещение организмов по земному шару, является температур а. Ряд животных может вырабатывать постоянную внутреннюю температуру тела. Такие животные называются гомеотермными, в отличие от животных пойкилотермных, температура тела которых непостоянна и определяется температурой среды. Промежуточную группу составляют животные гетеротермные, которые в активном состоянии гомеотермны, а при впадении в продолжительную спячку, длящуюся иногда до полугода и более, теряют способность к терморегуляции. В этот период число вдохов и выдохов у них составляет 1—2 в минуту, с такой же скоростью сокращается сердце.
Абсолютной грани между пойкилотермными и гомеотермными животными не существует. Многие пойкилотермные животные могут при движении продуцировать энергию, что приводит к подъему температуры их тела иногда значительно выше температуры среды. Так, температура тела многих насекомых (шмели, крупные бабочки) во время их полета может превышать температуру среды на 10—15°. Точно так же и некоторые пойкилотермные водные животные в состоянии продуцировать значительные количества тепла, несмотря на высокую скорость теплоотдачи в водной среде. Например, рыба тунец при быстром движении может иметь температуру тела, превышающую на 8—9° температуру воды.
С другой стороны, те птенцы, которые выходят из яйца неспособными к самостоятельной жизни (у так называемых птенцовых птиц), имеют не вполне постоянную температуру тела: она падает, когда родители не греют их своим телом, и поднимается, когда родители находятся на гнезде. Температура слепых голых детенышей обыкновенной полевки может падать до 8—9 и даже 4—5°, а при согревании остывших зверьков вновь поднимается до нормальной для них температуры 28—30°. Появление способности к терморегуляции совпадает у детенышей с их прозреванием.
Температура тела взрослых полевок и некоторых других мелких грызунов колеблется в пределах 28—41° в зависимости от условий, в которых зверек обитает, и от состояния зверька (так, зверек с намокшей шкуркой быстро охлаждается). При понижении температуры тела ниже 28° зверек быстро погибает. Поэтому для них не опасны летние дожди, которые не вызывают переохлаждения животных, но при осенних дождях зверькам грозит гибель в результате понижения температуры тела ниже минимальной. Колебания температуры тела отмечены также для колибри, стрижей и некоторых других птиц.
Гомеотермность («теплокровность») — существенное преимущество животного в борьбе за существование, именно поэтому гомеотермные животные — млекопитающие и птицы — по сравнению с пойкилотермными наиболее широко расселены по Земле.
Все организмы, населяющие нашу планету, по отношению к температуре среды могут быть эвритермными (обитающими при значительных колебаниях этого фактора) и стенотермными (обитающими при узких колебаниях температур). Некоторые стенотермные организмы обитают только при высоких температурах. Это теплолюбивые формы. Часть из них может обитать в теплых источниках с температурой, равной 70—80°. В таких условиях живут представители сине-зеленых водорослей и бактерий. Ряд животных и некоторые водоросли обитают в источниках с температурой 50—60°. Обычно белки свертываются при 40—45°. Непонятно поэтому, какие особенности позволяют этим организмам выживать при таких температурах. Другие стенотермные организмы являются холодолюбивыми. Стенотермными могут быть не только пойкилотермные, но и гомеотермные животные. Так, обитатели тропических широт — носороги, слоны, полуобезьяны, большая часть обезьян, попугаи — теплолюбивы; белый медведь, пингвины и другие обитатели полярных стран — холодолюбивы.
Температурный режим на суше изменяется в двух направлениях: средние годовые температуры воздуха понижаются от тропиков к полярным широтам, амплитуды суточных и годовых температур возрастают от окраин в глубь континентов. Любая точка земного шара характеризуется определенными средними суточными, месячными и годовыми амплитудами температур, определенной продолжительностью и температурным режимом различных времен года. Эти особенности температурного режима ограничивают возможности существования организмов в том или ином месте.
Растительные сообщества оказывают существенное влияние на годовой и суточный ход температур. Температура почвы на глубине 10 см имеет наиболее ровный ход и несколько ниже под ненарушенным луговым сообществом, несколько выше и характеризуется более значительными колебаниями под лугом с удаленным моховым покровом. Наконец, более высокие температуры с заметными колебаниями свойственны участкам луга, на которых был удален и моховой, и травяной покров. Еще более сильно влияют на температурный режим леса. Под пологом леса уменьшаются суточные и сезонные амплитуды температур, а средние суточные и годовые температуры ниже, чем на открытом месте.
Развитие растений и пойкилотермных животных находится в прямой зависимости от температуры среды. Оно начинается с той температуры, которая для данного вида является биологическим нулем (ниже которой он развиваться не может). Далее, до достижения оптимальной температуры повышение температуры ускоряет развитие. Поэтому в экологии принято вычислять суммы эффективных температур, необходимых для прохождения тех или иных стадий развития.
Обычный метод подсчета количества тепла, необходимого для жизнедеятельности организма или для выполнения им каких-либо функций, — метод подсчета суммы эффективных температур. В качестве нижней точки отсчета берут обычно тот минимум, при котором начинается развитие организма, или какой-либо условный рубеж (0°, +5°, + 10°). Зная на основе опытов, какая сумма тепла необходима для развития организма (например, для созревания плодов, зацветания, превращения личинки в куколку, вылета из куколки взрослого насекомого и т. д.), определяем по формуле, какое число дней необходимо для выполнения этой функции 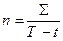
где п — искомое число дней, Σ — сумма тепла, необходимая для развития данного организма или выполнения данной функции, Т — постоянная температура периода развития данного организма, t — нижний температурный порог развития или избранный нами условный температурный рубеж. Если температура исследуемого нами периода непостоянна, то просто складывают средние суточные температуры до тех пор, пока полученная сумма не достигнет суммы тепла, необходимого для развития данного организма или для выполнения данной функции. День, когда это случилось, и будет последним из числа необходимых дней.
Однако определение суммы эффективных температур дает лишь общее представление о сроках наступления того или иного явления в жизни растения или животного: колебания температур в период развития, переходящие за нижний порог развития, могут заметно нарушить указанную выше картину. В этих случаях приходится вносить поправки.
При оценке влияния температурного режима различных регионов на гомеотермных животных применяют два правила — Бергмана и Аллена. Не следует только абсолютизировать эти правила. Применение этих правил основано на том, что гомеотермные животные производят тепло всей массой своего тела, и отдают его своей поверхностью. При увеличении размеров животного отношение величины объема к поверхности становится в этом отношении более благоприятным. В этом нас может убедить простой пример. Кубик с длиной ребра в 1 см имеет объем 1 см3 и поверхность 6 см2. Если длину ребра увеличить вдвое (до 2 см), то объем такого кубика будет равен 8 см3, а поверхность — 24 см2. Таким образом, при увеличении линейных размеров вдвое отношение объема к поверхности изменится с 6 до 3, т. е. станет более благоприятным с точки зрения соотношения между производством тепла и его отдачей. Из этого следует, что в более холодных районах для животного одного и того же вида более, благоприятны крупные размеры. Правило Бергмана и гласит о том, что в более северных районах нашего полушария размеры животных одного и того же вида становятся более крупными. Существенным дополнением к этому правилу служит правило Аллена, согласно которому у животных одного и того же вида, обитающих в более северных районах, размеры выступающих из общего абриса частей тела (уши, хвост, лапы) становятся более мелкими.
Не меньшее влияние, чем температура, оказывает на общие черты распределения организмов влажность. Имеются организмы эвригигробионтные, могущие существовать при самых различных значениях влажности: ксерофильные — обитающие в засушливых местообитаниях, мезофильные — в условиях среднего увлажнения, гигрофильные — при высоких значениях влажности. При этом способы приспособления организмов к жизни в одних и тех же условиях увлажнения разнообразны. Так, в условиях значительной сухости климата одни растения как бы «убегают» от засухи, заканчивая развитие в тот период, когда засуха еще не ощущается. Однолетние растения-эфемеры успевают развиться из семени, вырасти, отцвести и принести плоды, а затем отмереть за тот непродолжительный период, когда в аридных регионах влага (снеговая или дождевая) еще значительна. Многолетние растения-эфемероиды, развивающиеся из луковиц, клубней или корневищ, за тот же непродолжительный период приносят семена, но, в отличие от эфемеров, не отмирают полностью, а сохраняют на период засухи подземные запасающие органы — клубни, луковицы и корневища.
Другие виды, преимущественно водоросли, лишайники и мхи, обитающие в засушливых условиях — пойкилоксерофиты, — высыхают на период засухи так, что легко крошатся, будучи взятыми в руки, но при увлажнении вновь оживают и продолжают ассимилировать. Некоторые растения запасают влагу на сухой период. Это суккуленты. Они накапливают влагу в стеблях (стеблевые суккуленты) или в листьях (листовые суккуленты). Примерами первых могут служить кактусы, кактусовидные молочаи; вторых — алоэ, агавы, очитки и др. Сочные стебли и листья суккулентов плохо переносят морозы, поэтому они встречаются преимущественно там, где холодного периода нет или он кратковремен. Эти растения крайне медленно расходуют влагу: у них мало устьиц, которые при недостатке влаги очень плотно закрываются. Для фотосинтеза этим растениям приходится использовать наряду с углекислотой воздуха и тот углекислый газ, который накапливается в межклетниках этих растений при дыхании. Поэтому растут эти растения очень медленно. Зато влагу они хранят настолько надежно, что сорванный суккулент остается в сухой комнате живым в течение нескольких лет (рис. 15).
Некоторые растения аридных районов обладают очень длинными корнями, которые могут достигать грунтовых вод даже на глубине 16— 18 м. Такие растения, живя в пустыне, находятся в условиях достаточного водоснабжения, не испытывая недостатка влаги.
Другие растения аридных районов резко сокращают транспирацию, имея свернутые в трубку узкие или мелкорассеченные листья, покрытые густым опушением, с малым числом устьиц. Эти виды не являются суккулентами, так как у них нет сочных, мясистых стеблей или листьев, но в приспособлении к жизни в сухих регионах они идут по тому же пути сокращения обмена со средой и экономии влаги. Однако благодаря ограниченности связей со средой эти растения очень медленно развиваются.
Настоящие ксерофиты имеют густую сеть жилок и значительное число устьиц. Клеточные стенки листовых покровов плотны, часто пропитаны восковым веществом (кутинизированы) или одеты волосками. Испарение сквозь клеточные стенки, помимо устьиц, у этих растений сведено к минимуму, а устьица обладают способностью быстро закрываться при недостатке влаги и столь же быстро открываться при ее достаточном количестве. Хорошо развитые механические ткани не позволяют им завядать при потере влаги. Плазма этих растений хорошо переносит длительное обезвоживание. Таким образом, эти растения приспособлены не столько к недостатку влаги, сколько к ее частым колебаниям.
Столь же разнообразны и приспособления животных к обитанию в аридных регионах.
Некоторые животные запасают влагу. В мочевом пузыре запасают воду черепахи Галапагосских островов и одна из австралийских пустынных жаб. Последняя в период дождей раздувается, как шар, а к концу сухого периода, израсходовав воду, делается плоской.
Другие животные, например многие виды грызунов и насекомых, довольствуются теми незначительными количествами влаги, которые они получают с пищей. Эту пищу составляют растения, особенно их более влажные подземные органы, а также некоторые беспозвоночные животные, составляющие пищу грызунов.
Третьи впадают в летнюю спячку, накопив перед этим значительные количества жира. Во время спячки, проводимой этими животными в глубоких норах (таковы, например, суслики), процессы их жизнедеятельности резко замедлены — сердце делает 1-2 удара в минуту, число вдохов и выдохов также составляет 1-2, поэтому запасов жира, распадающегося во время спячки на воду и углекислый газ, хватает зверькам для поддержания жизни.
Четвертые накапливают значительные запасы жира и расходуют его, не впадая в спячку, подолгу не получая ни пищи, ни питья. Так, за счет метаболического распада жиров и выделения в этом процессе воды восполняют недостаток во влаге верблюды.
Виды, обладающие быстрым бегом (антилопы) или полетом (рябки), могут посещать водоемы, находящиеся на значительном расстоянии от места их жительства. Иногда они возвращаются с водоемов с зобами, полными водой (рябки).
Наконец, многие беспозвоночные весь засушливый период проводят в виде цист, одетых плотными оболочками, или спор, а некоторые мигрируют на весь сухой период в более благоприятные по режиму влажности регионы. У обитателей влажных мест — гигрофилов — приспособления к обитанию в этих условиях не столь многообразны, чем у обитателей сухих местообитаний — ксерофилов. Это, видимо, объясняется тем, что влажные местообитания значительно менее экстремальны, чем сухие.
У растений, обитающих в таких условиях, как правило, отсутствуют приспособления против завядания и быстрой потери влаги, механическая ткань, как правило, развита слабо, сильно развиты воздухоносные полости в корнях, стеблях и листьях. У животных, связанных с такими местообитаниями, наблюдается высокий уровень водного обмена и отсутствуют приспособления к его регулированию.
Мезофильные растения и животные характеризуются чертами, промежуточными между гигро- и ксерофильными видами.
Правило Глогера, заключающееся в том, что подвиды одного и того же вида млекопитающих животных, обитающие в более аридных условиях, окрашены светлее подвидов, свойственных более гумидным районам, основано, видимо, на том, что более темные вещества, определяющие окраску шерсти (меланины), отлагаются интенсивнее в более влажных регионах.
Осадки, выпадающие над сушей, имеют различную судьбу: они могут образовывать поверхностный и внутрипочвенный сток, испаряться, транспирироваться растениями. На все перечисленные компоненты растительный покров оказывает то или иное влияние. Лес самим своим существованием способствует более интенсивному выпадению осадков, над открытыми пространствами их выпадает меньше. Доля осадков, задерживаемая кронами деревьев в лесу или травостоем на лугах, в прериях и саваннах, различна и зависит от особенностей растительного покрова. Так, в разных типах леса доля осадков, задерживаемых кронами, стекающих по стволам деревьев и проникающих сквозь кроны непосредственно к почве, различна (рис. 16). Влияние на задержание осадков кустарниковых и травянистых сообществ значительно слабее по сравнению с лесом, но выражено достаточно четко. В лесу возрастает относительная влажность воздуха по сравнению с открытыми участками, уменьшается испарение с поверхности почвы (чему способствует лесная подстилка). Однако к влаге, испаряющейся с поверхности почвы, в сообществах присоединяется и влага, испаряющаяся с поверхности стволов, стеблей и листьев. Лес может вернуть в атмосферу до 1/3 влаги, выпавшей над ним в виде осадков. В травянистых сообществах эта доля меньше.
Различия в испарении с поверхности почвы в разных сообществах показаны на рис. 17. Под лесом понижается уровень грунтовых вод (рис. 18).
Транспирация, производимая лесными растениями, значительно интенсивнее транспирации луговых трав. Общая сумма транспирируемой и испаряемой с поверхности почвы воды выглядит в лесу и на лугу следующим образом (табл. 3).
Для облесенных территорий характерно резкое замедление поверхностного стока, который в этих случаях не носит катастрофического характера (рис. 19).
Ветер играет существенную роль в распространении семян и плодов, спор растений и покоящихся зачатков животных. Плоды и семена растений могут иметь специальные приспособления, облегчающие этот перенос, повышающие парусность переносимых зачатков. Это либо летучки из более или менее многочисленных волосков, либо крылатки, пленчатые или кожистые. Иногда плодик заключен в пленчатый мешочек, что также облегчает его перенос. Не меньшее значение имеет ветер и для опыления цветков растений. Цветки ветроопыляемых растений обычно производят значительно большее количество пыльцы по сравнению с опыляемыми насекомыми, так как в первом случае шансов попасть на рыльце пестика у пылинок много меньше. Пылинки легкие, иногда с воздушными мешками, облегчающими их полет (например, у хвойных). Рыльца пестиков ветроопыляемых цветков высовываются из цветков и часто бывают перистыми, поэтому они имеют большую «ловчую поверхность». Ветроопыляемые растения, как правило, растут большими группами, многие деревья и кустарники с цветками, опыляемыми ветром, цветут до распускания листьев.
Таблица 3
Годовой расход воды в лесу и на лугу (в мм) (по В. Н. Оболенскому, 1935)
| Растительность | Испарение с поверхности почвы | Транспирация | Всего |
| Лес | |||
| Луг |
Одновременно ветер оказывает формообразующее влияние на растения. Он может ломать ветви деревьев, а ветер ураганной силы ломает у основания стволы деревьев или вырывает их с корнем. Так, в лесах влажных тропиков образуются прогалины на месте вырванных ураганом деревьев. Обширные ветровальные участки наблюдаются в лесах умеренного пояса, преимущественно хвойных, особенно еловых, поскольку корни ели не идут глубоко в землю, а простираются близко к поверхности почвы. Ветер способствует образованию флагообразных форм деревьев, особенно у верхней границы их произрастания: в горах и у пределов леса на севере. Несомненно, что ветер играет существенную роль в безлесии тундр. Механизм этого влияния оценивается разными авторами различно. А. О. Чильман полагает, что зимой, когда почва промерзает, а ветви, не укутанные снегом, продолжают транспирировать, все части дерева, поднимающиеся над поверхностью снега, высыхают. Б. Н. Городков считает, что наиболее критическим периодом в этом отношении оказывается весна, когда ветви уже сильно прогреваются солнцем, а почва с корнями еще не оттаяла. Какой из этих точек зрения не придерживаться — значение ветра для безлесья тундры несомненно.
Прохождение северо-восточных границ ареалов широколиственных пород, свойственных европейской части Союза (липа мелколистная встречается к северо-востоку только до Тобольска, а восточные границы ареалов дуба обыкновенного и клена платановидного не доходят до Урала), связано с тем, что ветви этих пород зимой транспирируют значительно больше влаги на единицу поверхности по сравнению с хвойными, а низкие зимние температуры при большой скорости ветра не препятствуют зимней транспирации. В опытах известного русского ботаника А. Я. Гордягина транспирация с единицы поверхности ветвей дуба обыкновенного и других широколиственных пород в 2 раза и более превышает транспирацию с ветвей лиственницы. При этом транспирация ветвей дуба не прекращалась даже при значительных морозах: средние суточные потери веса у этого вида составляли при температурах 22, -8 и -18,4° соответственно 0,8, 0,6 и 0,2% живого веса. А. Я. Гордягин указывает, что в марте такие потери влаги не компенсируются за счет притока воды из более старых частей дерева. Таким образом, интенсивное зимнее и ранневесеннее испарение однолетними ветвями дуба и других широколиственных пород является одной из причин, препятствующих их распространению в области с более суровым климатом (на восток и северо-восток). Роль ветра при этом существенна.
При заселении островов ветер играет двойственную роль. Во-первых, он способствует переносу на острова диаспор (семян, плодов и спор) растений, а также различных крылатых беспозвоночных животных. Одновременно ветер уносит с островов и с побережий материков различные не очень хорошо летающие, хотя и крылатые формы. В результате отбора в таких местообитаниях многие виды, имеющие летающих крылатых родичей, становятся бескрылыми или, если сохраняют крылья, то приобретают привычку затаиваться под корой, среди листвы и т. п. Таким образом, ветер производит отбор среди обитателей участков, подверженных действию ветра, способствуя развитию бескрылых форм.
Влияние лесных сообществ на скорость ветра велико. Скорость ветра, дующего в сторону леса, уменьшается уже на некотором расстоянии от опушки. В лесу же она падает быстро (рис. 20). Аналогичное влияние, хотя и не столь сильное, оказывают на скорость ветра и другие растительные сообщества.
Газовый состав атмосферы в природе резко меняется только вокруг вулканов, где возрастает содержание газов, вредных для жизни животных и растений. В высокогорьях уменьшается общее давление атмосферы и парциальное давление кислорода. У животных способность приспосабливаться к пониженному содержанию кислорода сопряжена с увеличением содержания гемоглобина в крови. Те виды, которые обладают значительной пластичностью этого признака, могут обитать в горах выше животных, у которых содержание гемоглобина в крови меняется в узких пределах. У растений возможность существовать в высокогорьях связана с повышенным содержанием хлорофилла в тканях листа, однако механизм этого влияния не вполне ясен. Для оценки и наглядного представления о влиянии основных параметров климата на жизнь организмов применяется метод климограмм (климатограмм), при котором на оси координат наносятся основные климатические факторы (температура и влажность воздуха или температура и осадки). Имеются различные способы построения климограмм (рис. 21). Особенно детально разработаны способы построения климограмм немецким ботаником Г. Вальтером (рис. 22). На них видно не только соотношение температур и осадков, но также продолжительность сухого и влажного периодов и некоторые другие показатели.








