 2015-05-06
2015-05-06 338
338Система Х и У хромосом позволяет поддерживать равенство полов в популяции. При детерминации пола цветков у растений с Х и У хромосомами, растения с пестичными цветками, как правило гомогаметны (ХХ), а с тычиночными гетерогаметны (ХУ).
В популяции раздельнополых растений (конопля, спаржа, шпинат и др.) скрещиваются растения противоположных фенотипов (диссортативное скрещивание).
У растений с тычиночными цветками (ХУ) частоты аллелей и генотипов совпадают, если рассматриваемые гены локализованы в Х хромосоме (аллели генов находятся в гемизиготном состоянии):
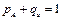
Для растений с пестичными цветками (ХХ) частоты генов и генотипов записываются как обычно (каждый ген в Х хромосоме имеет по два аллеля):
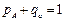
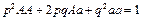
При случайном переносе пыльцы и случайном оплодотворении частоты генов и генотипов находятся в равновесии, т.е. закон Харди-Вайнберга выполняется и для генов, сцепленных с половыми хромосомами.
Однако, в случае сцепления с полом пестичные растения несут две дозы генов, а тычиночные – одну, поэтому равновесие частот генов и генотипов достигается только после нескольких поколений свободного размножения.
|
|
|
Приближение к равновесию носит колебательный характер. Это связано с тем, что тычиночные растения получают Х хромосому от пестичного растения, а пестичные – от обоих родительских растений.
Равновесная частота аллеля (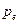 ) зависит от исходных частот в мужской (
) зависит от исходных частот в мужской (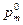 ) и в женской
) и в женской
(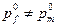 ) частях популяции следующим образом:
) частях популяции следующим образом:
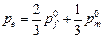
Пример
Популяция состоит из мужских и женских особей с контрастными частотами аллеля А:
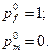
В результате первого свободного скрещивания
А//А х а/ → А//а + А/
генетическая структура женской части популяции потомства определяется в равной степени частотами аллелей в обеих исходных частях:
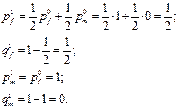
После второго свободного скрещивания:
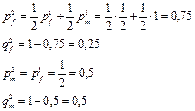
И тд.








