 2015-05-26
2015-05-26 5746
5746Тело первых вышедших на сушу растений ещё не было расчленено на побеги и корни. Оно состояло из ответвлений, одни из которых поднимались вертикально, а другие прижимались к почве и поглощали воду и питательные вещества. Несмотря на примитивное строение, эти растения были обеспечены водой и питательными веществами, так как имели небольшие размеры и жили около воды.
В ходе дальнейшей эволюции некоторые ответвления стали углубляться в почву и дали начало корням, приспособленным к более совершенному почвенному питанию. Это сопровождалось глубокой перестройкой их структуры и появлением специализированных тканей. Образование корней было крупным эволюционным достижением, благодаря которому растения смогли осваивать более сухие почвы и образовывать крупные побеги, поднятые вверх к свету. Например, у моховидных настоящих корней нет, их вегетативное тело небольших размеров — до 30 см, обитают мхи во влажных местах. У папоротникообразных появляются настоящие корни, это приводит к увеличению размеров вегетативного тела и к расцвету этой группы вкаменноугольный период.
Типы корневых систем
§ В стержневой корневой системе главный корень сильно развит и хорошо заметен среди других корней (характерна для двудольных).
§ В мочковатой корневой системе на ранних этапах развития главный корень, образованный зародышевым корешком, отмирает, а корневая система составляется придаточными корнями (характерна для однодольных). Стержневая корневая система проникает в почву обычно глубже, чем мочковатая, однако мочковатая корневая система лучше оплетает прилегающие частицы грунта.
§ Придаточные корни растут непосредственно из стебля. Они отрастают от луковицы (представляющей собой особый стебель) или от садовых черенков.
§ Воздушные корни. Корни, которые растут от стебля. но не проникают в землю. Они используются лазящими растениями для закрепления, как например, у плюща.
§ Опорные (ходульные) корни. особый тип воздушных корней. Они отрастают от стебля и затем проникают в землю, которая может быть покрыта водой. Они поддерживают тяжёлые растения, например мангры.
18. Типичный корень представляет собой подземный орган, присущий всем высшим растениям (кроме мхов). Корень служит для закрепления растения в почве, поглощения из почвы воды с растворенными в ней солями, в корне часто откладываются запасные продукты, корень участвует в синтезе органических веществ, служит для вегетативного размножения. Корень никогда не несет на себе листьев, поэтому по сравнению с внутренней структурой стебля у корня она относительно проста.
Корень по длине можно разделить на несколько участков, имеющих различное строение и выполняющих различные функции. Эти участки называют зонами корня. Выделяют корневой чехлик и следующие зоны: деления, растяжения, всасывания и проведения.
Дифференциация тканей корня происходит в зоне всасывания. По происхождению это первичные ткани, так как они образуются из первичной меристемы конуса нарастания. Поэтому микроскопическое строение корня в зоне всасывания называют первичным. У однодольных растений первичное строение сохраняется и в зоне проведения. Здесь лишь отсутствует самый поверхностный слой с корневыми волосками - ризодерма (эпиблема). Защитную функцию выполняет ниже лежащая ткань - экзодерма.
В первичном строении корня выделяют три части: ризодерму, первичную кору и осевой (центральный) цилиндр.
Строение ризодермы рассматривалось в теме "Покровные ткани".
На первичную кору приходится основная масса первичных тканей корня. Ее клетки накапливают крахмал и другие вещества. Эта ткань содержит многочисленные межклетники, имеющие значение для аэрации клеток корня. Наружные клетки первичной коры, лежащие непосредственно под ризодермой, называются экзодермой. Основная масса коры (мезодерма) образована паренхимными клетками. Самый внутренний слой носит название эндодермы. Это ряд плотно сомкнутых клеток (без межклетников).
Центральный или осевой цилиндр (стела) состоит из проводящих тканей, окруженных одним или несколькими слоями клеток - перициклом.
Внутренняя часть центрального цилиндра у большинства растений занимает сплошной тяж первичной ксилемы, дающий к перициклу выступы в виде ребер. Между ними размещаются тяжи первичной флоэмы.
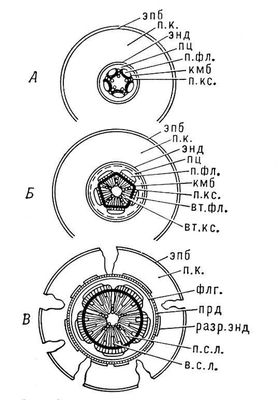
У двудольных и голосеменных растений уже в раннем возрасте в центральном цилиндре корня между ксилемой и флоэмой появляется камбий, деятельность которого приводит к вторичным изменениям и в конечном итоге формируется вторичная структура корня. К центру камбий откладывает клетки вторичной ксилемы, а к периферии - клетки вторичной флоэмы. В результате деятельности камбия первичная флоэма оттесняется кнаружи, а первичная ксилема остается в центре корня.
Вслед за изменениями в центральном цилиндре корня происходят изменения в коровой части. Клетки перицикла начинают делиться по всей окружности, в результате чего возникает слой клеток вторичной меристемы - феллогена (пробкового камбия). Феллоген, в свою очередь, делясь, откладывает наружу феллему, а внутрь - феллодерму. Образуется перидерма, пробковый слой которой изолирует первичную кору от центрального цилиндра. В результате вся первичная кора отмирает и постепенно сбрасывается; наружным слоем корня становится перидерма. Клетки феллодермы и остатки перицикла в дальнейшем разрастаются и составляют паренхимную зону, которую называют вторичной корой корня (рис. 70).
При развитии запасающей паренхимы главного корня происходит формирование запасающих корней или корнеплодов. Различают корнеплоды:
1. Монокамбиальные (редька, морковь) - закладывается только один слой камбия, а запасные вещества могут накапливаться либо в паренхиме ксилемы (ксилемный тип - редька), либо в паренхиме флоэмы (флоэмный тип - морковь);
2. Поликамбиальные - через определенные промежутки времени происходит заложение нового слоя камбия (свекла).
17. Корневой чехлик - защитное образование растущего кончика корня. Служит защитой преимущественно от механических повреждений. Это небольшой конусовидный колпачок, длиной обычно около 0,2 мм, реже, у воздушных корней, до нескольких миллиметров, прикрывающий нежные клетки апикальной меристемы кончика корня и часть его зоны роста.
Клетки корневого чехлика образуются при первичном росте корня за счет деления клеток части апикальной меристемы — калиптрогена.
Клетки корневого чехлика живут всего несколько суток и постоянно слущиваются с его поверхности. Наружные слои клеток чехлика секретируют слизь, состоящую в основном из полисахаридов. К этой слизи прилипают частицы почвы, что, возможно, защищает кончик корня от высыхания. Во многих источниках указывается также, что слизь служит смазкой при продвижении корня в почве.
Амилопласты центральной зоны корневого чехлика служат статолитами, которые обеспечивают георецепцию и положительный геотропизм растущего корня.
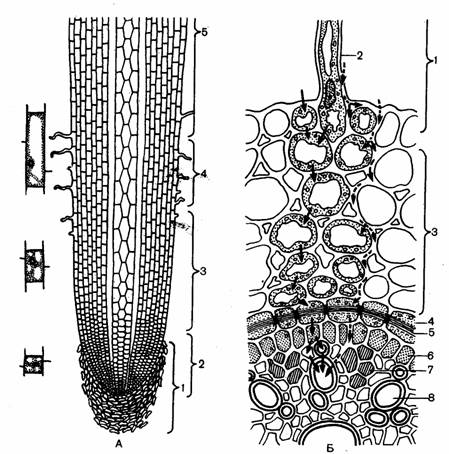
(А – продольный разрез: 1 – корневой чехлик; 2 – меристема; 3 – зона растяжения; 4 – зона корневых волосков; 5 – зона ветвления; Б- поперечный разрез: 1 –ризодерма; 2 – корневой волосок, 3 – паренхима, 4 – эндодерма; 5 – пояски Каспари, 6 – перицикл, 7 – флоэма, 8 – ксилема. Стрелки – пути передвижения веществ, поглощаемых из наружного раствора. Сплошные стрелки – путь раствора по симпласту; прерывистые – по апопласту.)
19. ВТОРИЧНОЕ УТОЛЩЕНИЕ, в ботанике - увеличение толщины стебля и корней, особенно у деревьев и кустарников, в результате образования новых клеток в КАМБИИ. Клетки камбия дифференцируются внутри - в элементы вторичной КСИЛЕМЫ, а снаружи - в клетки вторичной Флоэмы. Новый цилиндр, образованный камбием возле поверхности стебля или корня, может создать жесткий внешний слой клеток ЛУБА или КОРЫ.
(Схема вторичного утолщения корня: А — первичное строение; Б, В — вторичное утолщение; эпб — эпиблема, п. к. — первичная кора, энд — эндодерма, пц — перицикл, п. фл. — первичная флоэма, кмб — камбий, п. кс. — первичная ксилема, вт. фл. — вторичная флоэма, вт. кс. — вторичная ксилема, флг — феллоген, прд — перидерма, разр. энд. — разрушенная эндодерма, п. с. л. — первичный сердцевинный луч, в. с. л. — вторичный сердцевинный луч.)
20. Побе́г — один из основных вегетативных органов высших растений, состоящий из стебля с расположенными на нём листьями и почками.
Вегетативный невидоизменённый побег — единый орган растения, состоящий из стебля, листьев и почек, формирующийся из общего массива меристемы(конуса нарастания побега) и обладающие единой проводящей системой. Стебли и листья, являющиеся основными структурными элементами побега — часто рассматриваются как его составные органы, то есть органы второго порядка. Кроме того, обязательная принадлежность побега — почки. Главная внешняя черта, отличающая побег от корня — наличие листьев.
Для побега характерна метамерность, то есть чередование одинаковых частей — метамеров. Листья располагаются на стебле в определённом порядке, в соответствии с типом листорасположения, а участок стебля на уровне отхождения листа называется узлом. Если основание листа полностью окружает стебель, узел называется закрытым, в противном случае — открытым. Промежутки стебля между соседними узлами называют междоузлиями. Каждый повторяемый отрезок побега, который включает в себя узел с находящимся под ним междоузлием, а также расположенные на этом отрезке листья и почки называют метамером. Угол между листом и вышележащим междоузлием называют листовой пазухой, или пазухой листа.
Морфологическая классификация. можно назвать побег, состоящий из одного прироста (по не отдельные приросты одного побега!), элементарным или простым; побег, наросший в течение одного вегетационного периода,— годичным, побег, живущий несколько лет и состоящий из нескольких приростов,— многолетним или сложным. В.Н. Любименко (1900) подразделил побеги деревьев на три группы: 1) обыкновенные или длинные (с листьями срединной формации и удлиненными междоузлиями); 2) типичные укороченные (с листьями срединной формации и укороченными междоузлиями); 3) скрытые укороченные (с листьями низовой формации и укороченными меж доузлиями).
Дод (Dode, 1905) дает следующую классификацию побегов. А. Одно-трехлетние побеги: 1) ауксибласты — ростовые побеги, появляющиеся на концах молодых ветвей; 2) мезобласты — промежуточные побеги, характеризующиеся тонкостью и более мелкими листьями; 3) хилые побеги — обычно следующего порядка. Б. Годичные побеги старых пяти-шестилетних ветвей: 4) лампробласты — массивные побеги; 5) тениобласты — тонкие побеги, возникшие летом на коротких весенних побегах; 6) брахибласты — короткие побеги старых ветвей, которые имеют вид узловатых веточек, состоящих из мелких годичных приростов. Из сложного подразделения типов побегов Дода некоторые термины прочно вошли в морфологическую литературу, например брахибласты, противопоставляемые ростовым побегам, называемым ауксибластами.
Е. Варминг (Warming, 1908) делит побеги на длинночленистые (ассимиляционные) и короткочленистые (розеточные). Б. В. Закржевский и Е. П. Коровин (1935), И. И. Курындин и др. (1947), А. В. Гурский и др. (1953), А. А. Федоров и др. (1962) следуют в основном Доду (Dode, 1905), деля побеги на два типа: удлиненные (ауксибласты) и укороченные (брахибласты). К брахибластам эти авторы относят укороченные цветоносные побеги многих плодовых деревьев (например яблонь» именуемых в практике садоводства плодушками или кольчатками).
Л. В. Кудряшов и Р. П. Барыкина (1970) для стланиковых кустарников выделяют четыре типа побегов: удлиненные вегетативные, полурозеточные вегетативные, укороченные (розеточные вегетативные), укороченные генеративные.
Г. Н. Зайцев (1962) выделяет следующие типы побегов жимолостей в зависимости от направления роста и расположения почек, из которых они развиваются: ортотропные годичные побеги, возникающие на адаксиальной стороне материнского побега и направленные вверх; амфитропные побеги, возникающие из трансверсальных почек и направленные примерно горизонтально; ортоамфитропные побеги, образованные из адаксиальных и трансверсальных почек; изофилъные побеги, развивающиеся обычно равномерно из всех категорий почек и расположенные под большим углом к горизонту; гипотрофные побеги, направленные вниз.
Морфолого-биологическая классификация. И. Г. Серебряков (1949) делит побеги древесных растений на вегетативные (укороченные и удлиненные — ростовые) и цветоносные, или плодущие (как правило, укороченные). Р. А. Мастинская (1971) по сочетанию двух признаков выделяет побеги ежевики Rubus caesius: 1) вегетативно-ортотропные, 2) вегегативно-плагиотропные однолетние, 3) вегетативно-плагиотронные двухлетние, 4) генеративные побеги из пазушной почки дефинитивного листа, 5) генеративные прикорневищные побеги. Биологическая классификация И. Г. Серебряков (1949) в зависимости от специализации делит побеги на три категории: слабо, неполно и крайне специализированные.
Н. Е. Булыгин (1965) классифицирует генеративные побеги на: 1) генеративные побеги, после плодоношения полностью отмирающие: а) крайне специализированные, например у вяза Ulmus scabra Mill., б) специализированные, например у черемухи Padus racemosa (Lam.) Gilib.; 2) генеративно-ростовые, у которых отмирает генеративная часть: а) неполно специализированные, например у свидины Thelycrania аlbа (L). Pojark и клена маньчжурского Acer mandschurica Maxim, б) слабо специализированные, например у барбариса Berberis vulgaris L., в) неспециализированные, например у жимолости голубой Loniсera caеrulea L. По степени проявления вегетативной и репродуктивной функции Н. Е. Булыгин делит побеги на три основные группы: вегетативные, вегетативно-репродуктивные и строго репродуктивные.
У Е. Л. Нухимовского (1970) мы встречаем несколько подходов к разграничению побегов, например по характеру раз вития верхней части побега: 1) открытые, не заканчивающиеся соцветием, тип Loniсera caerulea; 2) условно закрытые, заканчивающиеся кистевидным соцветием, тип Padus racemosa; 3) закрытые, у которых верхушечные меристемы образуют закрытые соцветия, заканчивающиеся терминальным цветком, тип Rosa huntica. Е. Л. Нухимовский также классифицирует побеги по степени отмирания, предлагая понятие «резид». Резид — стеблевая часть побега, которая стала многолетней после завершения им фазы видимого или верхушечного роста. В зависимости от степени отмирания резиды делятся на максимо-, медио- и миниморезиды. Побеги, не образующие резидов, т. е. однолетние, эфемерны. Побеги, образующие медио- и миниморезиды, по аналогии с эфемерными можно назвать зфемероидными.
Шпет (Sp а t, 1912) делит побеги в зависимости от периода покоя и сформированности почек на: 1) настоящие ивановы, в которых после роста весенних побегов почка не сформирована, затем она формируется и дней через 40 начинает расти; 2) надставленные ивановы, которые возникают из терминальных конусов нарастания без образования закрытой почки и обычно без перерыва роста, редко после краткой его остановки; 3) с иллептические, в которых на побеге совсем не образуется верхушечная почка и верхушка непрерывно продуцирует листья, в пазухах закладываются почки, они сразу трогаются в рост одновременно с продолжающимся ростом «обычного годичного побега"; 4) пролептические, развивающиеся на годичном побеге из уже сформированных пазушных ночек, которые находились в состоянии покоя.
21. По́чка — в ботанике зачаток побега; обычно образуется у растения в пазухе листа (пазушная почка) либо на конце побега (верхушечная почка, или терминальная почка), либо на взрослых органах (стебле, листе, корне; в этом случае он не связан ни с верхушками побегов, ни с узлами и не обнаруживает чётких закономерностей в своем расположении) — придаточная почка. Однажды появившись, почка может оставаться нераскрытой на определённый промежуток времени в состоянии покоя (спящая почка), либо тут же из неё начинает развиваться побег. Вегетативные почки, из которых развиваются вегетативные побеги, состоят из зачаточного стебля и зачаточных листьев. Генеративные почки, из которых развиваются цветки или соцветия, состоят из зачатков соцветий и цветков.
Почки у многих древесных растений, особенно в умеренном или прохладном климате, покрыты видоизменёнными листьями, называемыми чешуями. Такие почки называют закрытыми, или защищёнными. Если чешуи отсутствуют, то такие почки называют голыми, или незащищёнными.
У кактусов имеются особые почки — ареолы, а почечные чешуи у них видоизменены в иглы. Иногда чешуи почек покрыты смолистым веществом для большей защиты. Когда почка раскрывается, чешуи могут увеличиться, но чаще опадают, оставляя на поверхности стебля ряд горизонтально-вытянутых рубцов. На основании количества таких рубцов можно частично определить возраст растения, так как в течение одного сезона растущий стебель образует на конце только одну почку, после которой образуется рубец. С течением времени такие рубцы постепенно стираются, так что определить полный возраст растения с помощью этого метода бывает затруднительно.
У голой почки слаборазвитые листья часто покрыты избыточным ворсом. Голые почки присутствуют у таких кустарников, как Сумах (Rhus) иКалина (Viburnum), а также у травянистых растений. У последних очень часто почки более мелкие и состоят из однообразной массы клеток в пазухе листа.
Поскольку почки формируются в пазухе листа, их распределение на стебле аналогично распределению листьев: они могут располагаться попеременно, супротивно, мутовчато, а также верхушечно на конце стебля. У некоторых видов растений почки могут появиться не в обычных местах их развития — такие почки называют придаточными. Например, у древесных растений придаточные почки могут развиться на корневой системе, в результате чего возникает корневая поросль. Некоторые травянистые растения, такие как, например, Татарник полевой (Cirsium setosum), переживают зиму за счёт придаточных почек, так как его главный стебель отмирает ещё в первый год.
Если внутри почки заключены только зачатки цветков (одного или нескольких), то её называют цветочной почкой. Цветочные почки противопоставляют листовым, заключающим в себе лишь зачатки обыкновенных листьев, и смешанным, в которых, кроме листьев, находятся и зачатки цветов.
Почка — это находящийся в состоянии относительного покоя зачаточный побег с очень сильно сокращенными междоузлиями. Морфологические признаки почек самые разнообразные.
По расположению на побеге различают:
верхушечные (концевые, терминальные) и боковые (латеральные) почки,
по месту формирования — пазушные (превентивные) и придаточные (адвентивные). Пазушные почки формируются в пазухах листьев, а придаточные могут образовываться эндогенно на различных участках стебля, корнях, а у отдельных культур и на листьях. Придаточные почки имеют большое значение при восстановлении растений после повреждений и при их вегетативном размножении.
На стебле почки располагаются, как правило, экзогенно (снаружи), но могут быть и эндогенные, защищенные перидермой стебля (актинидия).
В узле побега почки прикрепляются к листовой подушечке непосредственно и называются в таком случае сидячими. У смородины, вишни, груши встречаются черешковые почки, когда у основания почки имеется небольшое стеблевое образование в виде ножки.
В зависимости от длины междоузлия почки на стебле могут быть размещены сближено, скученно, расставлено, а иногда и супротивно, что совпадает с листорасположением.
Непосредственно в пазухе листа они могут располагаться одиночно или группами по 2...3 и более. Групповые почки могут иметь вертикальное (одна над другой) или горизонтальное (рядом друг с другом) размещение. В первом случае такие почки называют сериальными (малина, жимолость съедобная, грецкий орех), а во втором — коллатеральными (косточковые породы, миндаль).
По типу новообразований и выполняемых функций различают вегетативные, генеративные и вегетативно-генеративные почки. Последние две группы называют цветковыми почками.
У вегетативно-генеративных почек одновременно формируются полноценные зачатки генеративных и вегетативных органов, поэтому их называют смешанными цветковыми почками или просто смешанными почками. Такие почки характерны для семечковых пород и некоторых ягодных (смородина, крыжовник).
Почки различаются по размерам, внешнему виду и внутреннему строению. Обычно цветковые почки, особенно у семечковых пород, более крупные и более округлые, чем вегетативные, что хорошо заметно в конце вегетации растений. Эти различия дают возможность прогнозировать урожай будущего года и нормировать его в весенне-зимний период с помощью обрезки. Почки, расположенные в разных частях одного побега, морфологически неоднородны. Это свойство называют разнокачественностью почек и используют при отборе почек для прививки.
Сверху почки защищены почечными чешуями, причем их число бывает различным. По числу чешуи у облепихи определяют мужские и женские цветковые почки и соответственно пол растения. У земляники, калины, малины почечные чешуи отсутствуют, такие почки называют голыми.
Почка содержит в себе структурные элементы будущих побегов. Вегетативные почки состоят из оси, конуса роста, зачатков листьев с зачатками пазушных почек.
Смешанные цветковые почки вместо конуса роста имеют зачатки цветков, а генеративные почки — только зачатки цветков (иногда вместе с листьями). Сформировавшиеся на побеге почки прорастают в новые образования в различное время в зависимости от породно-сортовых особенностей и природных условий. Если в типичных условиях они прорастают в текущий вегетационный период, то их называют скороспелыми (чаще у косточковых), а если в начале следующего года — позднеспелыми (у семечковых). Только из части проросших почек могут развиться побеги ростового типа (длиной более 10...20 см).
Почки, способные к прорастанию в текущем году, считаются скороспелыми. Такие вегетативные почки встречаются у косточковых пород, особенно у персика, абрикоса, отличающихся загущенной кроной.
Скороспелостью могут обладать и цветковые почки, но, как правило, они имеются только у ремонтантных сортов земляники, малины. Позднеспелые почки прорастают на следующий год после того, как сформируются, они характерны для семечковых пород.
Часть почек на побегах, особенно у основания, плохо развиваются и не прорастают в следующем году или в течение нескольких лет. Такие почки считаются спящими. Оставаясь внешне недеятельными, они сохраняют поверхностное расположение за счет удлинения своей оси в соответствии с увеличением диаметра ветви. Этот ежегодный прирост спящих почек прослеживается по так называемым почечным следам на продольных срезах ветви. Способность к прорастанию этих почек сохраняется у многолетних растений в течение нескольких десятков лет и определяет их побеговосстановительную способность.

