 2015-05-30
2015-05-30 4541
4541Жизнь насекомых в лесу обусловлена многообразными, часто очень сложными, а порой и противоречивыми связями с другими организмами и между собой. Их можно разделить на межвидовые и внутривидовые отношения.
Межвидовые отношения. У насекомых межвидовые отношения обычно проявляются в виде симбиоза, паразитизма, хищничества и конкуренции.
Симбиоз— это устойчивые, разного свойства тесные взаимосвязи организмов различных видов, выгодные одному или обоим видам. Выделяют два типа симбиоза: мутуализм, при котором оба организма извлекают пользу от совместного существования; комменсализм, при котором взаимоотношения для одного вида полезны, а для другого большей частью нейтральны. Оба типа симбиоза широко распространены в природе. Так, муравьи питаются сахаристыми выделениями тлей и кокциД (некоторые виды чернецов), очищают при этом их колонии от клейких экскрементов и защищают от врагов (хищников и паразитов). Многие насекомые, питающиеся древесиной, заносят в ходы грибы, которые там развиваются и способствуют разложению клетчатки на составные химические соединения, которыми питаются насекомые—ксилофаги. Существуют мутуалисти-ческие отношения между термитами и обитающими в их кишечнике одноклеточными организмами.
Особенно многочисленны примеры комменсализма. Так, например, ряд насекомых прикрепляется друг к другу для передвижения (форрезия). Иногда насекомые одного вида живут за счет запасов другого, не причиняя ему существенного вреда (нахлебничество). Многие насекомые укрываются в гнездах муравьев, термитов, златогузки.
Паразитизм — это односторонне выгодное использование одним организмом другого организма в качестве источника пищи и среды обитания на протяжении большей или меньшей части своего жизненного цикла. Существуют первичные паразиты, которые в качестве хозяев используют фитофагов, и вторичные, или гиперпаразиты (сверхпаразиты); они используют в качестве хозяев других паразитов. В одном хозяине может развиваться одна или несколько особей паразита. Бывают случаи, когда одного хозяина заражает несколько видов паразитов (мультипаразитизм}. Формы паразитизма очень разнообразны..Среди насекомых существует много видов самых разнообразных паразитов. Большинство из них относится к отрядам перепончатокрылых и двукрылых (см. гл. XI).
Хищничество отличается от паразитизма в основном тем, что жертва сразу погибает от нападающего на нее хищника. При этом один хищник может уничтожить много жертв. В контакте с каждой жертвой он проводит ограниченное время. Хищные насекомые охотятся за гусеницами бабочек, истребляют личинок жуков, различных куколок и даже взрослых насекомых. Они ловят свою жертву, убивают ее и затем поедают. Типичные хищники: жужелицы, рыжие лесные муравьи, мухи-ктыри, песчаные осы и другие насекомые. Хищниками являются также насекомоядные птицы, летучие мыши, бурозубки и другие представители млекопитающих из отряда насекомоядных.
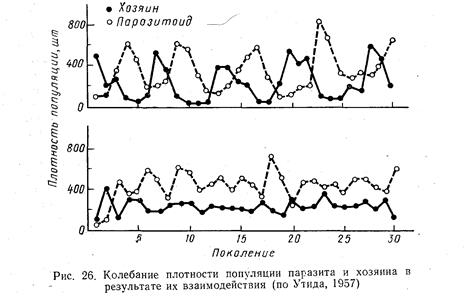
Численность хищника (паразита) и численность его жертвы (хозяина) тесно связаны между собой. Некоторые исследователи доказали, что при неизменных условиях численность жертвы увеличивается постепенно в течение ряда поколений, а затем падает; то же самое происходит с численностью хищника, но с некоторым запаздыванием. Когда жертва имеется в изобилии, то численность хищника возрастает, что в конечном итоге приводит к сокращению численности жертвы и в свою очередь вызывает уменьшение хищника, после чего численность жертвы может снова возрастать и цикл повторяется. На рис. 26 графически изображены колебания численности одного и того же вида хозяина (фасолевая зерновка), преследуемого медленно (вверху) и быстро (внизу) воспроизводящимся паразитом. При быстром размножении паразита он стабилизирует популяцию жертвы.\
Конкуренция между разными видами насекомых возникает в том случае, когда сходные потребности к условиям жизни полностью не удовлетворяются. Так, если на дубе одновременно развивается большое число гусениц нескольких видов чешуекрылых, питающихся листьями, и этих листьев всем не хватает, между гусеницами складываются конкурентные отношения. Очень часто популяция вида, чьи гусеницы отрождаются в момент распускания листьев, вытесняют гусениц других видов, появляющихся позднее, так как им уже не остается пищи.
Очень ярко проявляется межвидовая конкуренция у хищных насекомых. Например, муравей черный лазиус (Lasius niger} всегда вытесняет с занятой им территории желтого лазиуса (Lasius flavus). Конкурентные отношения между видами оказывают большое влияние на характер размещения и перемещения насекомых по территории.
Внутривидовые отношения. Особи в популяциях одного вида насекомого связаны различными по характеру и значению взаимоотношениями.
Отдельные особи и группы особей одного и того же вида могут воздействовать друг на друга прямо и активно или же косвенно, через изменение окружающей среды. Результаты таких отношении для отдельно взятой особи или группы особей могут быть весьма различны. Они могут стимулировать протекание жизненных процессов (рост, размножение, развитие) или ослаблять их. На большинство форм внутривидовые отношений определенное влияние оказывает численность участвующих в них особей. Регулирование численности достигается рядом внутривидовых механизмов. Типичным внутривидовым приспособлением является переход саранчи от одиночного к стадному образу жизни и последующие миграции стай. Гусеницы многих бабочек, питающиеся листвой древесных пород, живут в первых возрастах обществами, а затем расходятся и переходят к одиночному образу жизни, избегая таким образом конкуренции за пищу. В период же коллективного питания гусениц "интен-сивность последнего стимулировалась специфической средой их совместного обитания.
Характерным внутривидовым приспособлением служат гнезда у пушистого и кольчатого коконопрядов. В этих гнездах живет по нескольку сотен гусениц. Ночью температура гнезда на 8—12° С выше, чем окружающего воздуха. Это позволяет гусеницам легко перенести низкие ночные температуры.
Существует связь между размерами листа тополя и количеством откладываемых на него яиц тополевой молью. С увеличением размера листа возрастает и число яиц. Автоматическое регулирование численности происходит также у щитовок. При очень густом поселении одни особи вытесняют механически другие. Вытесненные особи не получают питания и погибают или голодают и становятся бесплодными.
При очень большой плотности популяции обычно возникает острая конкуренция между особями за пищу. В результате резко возрастает смертность, развиваются эпизоотии и численность автоматически сокращается, вновь увеличиваясь через определенный промежуток времени.
НАСЕКОМЫЕ КАК ЭЛЕМЕНТ ЭКОСИСТЕМЫ
Выше говорилось, что первичной элементарной структурной единицей биосферы является экосистема. Обычно ставят знак равенства между экосистемой и биогеоценозом. Однако есть некоторые расхождения между пониманием экосистемы в зарубежных работах и трудах В.Н. Сукачева (автора учения о биогеоценозах) и его школы.
В отличие от понятия экосистемы у зарубежных авторов, согласно которому экосистема может охватывать пространство любой протяженности — от капли прудовой воды и до Вселенной, — биогеоце-ноз имеет строго определенный объем. Биогеоценозы (экосистемы) являются частями земной или водной поверхности, однородной по топографическим, микроклиматическим, почвенным, гидрологическим и биотическим условиям..
В современной экологической литературе применяется термин экосистема. Он краткий и международный. Этот термина будет использован в дальнейшем.
Экосистема относительно, устойчива во времени и термодинамически открыта в отношении притока и оттока веществ в атмосферу.
Экосистемы сложились в процессе длительной эволюции, они являются слаженными, устойчивыми механизмами, способными путем саморегулирования противостоять как изменениям в среде, так и изменению в численности организмов.
Все взаимодействия компонентов экосистемы основаны в конечном счете на обмене веществом и энергией между ними. Основным источником энергии в экосистемах Земли служит Солнце. На Земле энергия солнечного излучения включается в биологический круговорот только через фотосинтез, осуществляемый автотрофными организмами. Кванты видимого света, проходя через экосистему, поглощаются хлорофиллом зеленых клеток растений. В процессе фотосинтеза автотрофные растения поглощают из атмосферы углекислый газ, из почвы—воду и минеральные вещества и с помощью энергии Солнца создают из них сложные органические соединения (углеводы, белки, жиры, органические кислоты и др.). Эти вещества богаты энергией и способны к различным химическим превращениям как в самих растениях, так и в телах различных гетеро-трофов, использующих энергию растительных материалов в процессе питания на свои жизненные функции.
Количество солнечной энергии, поступающей в экосистемы, ничтожно по сравнению с общим энергетическим бюджетом Земли. Всего около 0,1% энергии, получаемой от Солнца, связывается в процессе фотосинтеза. Однако за счет этой энергии может синтезироваться несколько тысяч граммов сухого органического вещества на 1 м2 в год. Более половины энергии, связанной при фотосинтезе, тут же расходуется в процессе дыхания самих растений. Часть же ее переносится через посредство ряда организмов по пищевым, или трофическим цепям экосистемы.
Пищевые цепи возникают как результат сложных пищевых взаимоотношений между растительными и животными организмами. Они объединяют прямо или косвенно большую группу организмов в единый комплекс.
Цепь питания обычно состоит из нескольких звеньев. Первое звено образуют продуценты, или производители. Это автотрофные растения. Они создают первичную биологическую продукцию и аккумулируют солнечную энергию.
Второе звено представлено консументами (консумцио—потребляю), или потребителями,— гетеротрофными организмами. Различают консументы первого порядка — фитофаги, второго порядка — плотоядные животные, третьего порядка — хищники,, питающиеся другими животными, и т. д.
Наряду с консументами существуют еще редуценты (редуцирую—уменьшаю), или деструкторы, которые разрушают и поедают мертвое органическое вещество и минерализуют его до простых неорганических соединений. К ним относится большая группа сап-рофагов, в которую входят грибы, микроорганизмы, многие членистоногие и др.
В каждой цепи формируются определенные трофические уровни, характеризующиеся различной интенсивностью протекания потокаизменениям, получившим название сукцессии. Если первый этап охватывает период жизни деревьев от появления их из семян до смыкания кронами и образования древостоя, второй наступает после смыкания деревьев кронами и длится до окончания интенсивного изреживания, третий этап-—это период средневозрастных и спелых насаждений, а четвертый характеризуется старением древостоя. Каждому периоду свойственны определенные энтомокомплексы дендрофильных насекомых (Воронцов, 1963).
По мере разрушения древостоя под влиянием самых различных причин (пожары, рекреационное воздействие, ветровалы и т. д.) происходит заселение деревьев насекомыми и грибами, комплексы которых последовательно сменяются под влиянием изменения физических свойств древесины. Наблюдается типичная сукцессия, хорошо изученная на многих древесных породах Б. М. Мамаевым и сотрудниками (рис. 31).
ДИНАМИКА ЧИСЛЕННОСТИ ПОПУЛЯЦИЙ
В экологии насекомых центральное место занимает динамика численности популяций. Плотность определяется числом особей, приходящихся на единицу площади или объема. Методы определения плотности популяции разнообразны и зависят от экологии особей данного вида и тех биотопов, которые она населяет. Характер распределения особей, составляющих популяцию, в пространстве может быть равномерным, случайным и скученным. Знание типа распределения особей в популяции важно при оценке ее плотности методом выборки.
Возрастной состав популяции имеет очень большое значение для ее существования и процветания. При благоприятных условиях в популяции присутствуют все возрастные группы и поддерживается более или менее стабильный состав. В быстро растущих популяциях доминируют интенсивно размножающиеся молодые особи, а в сокращающихся—старые, уже неспособные интенсивно размножаться. Такие популяции малопродуктивны, недостаточно устойчивы. На возрастной состав популяции большое влияние оказывает продолжительность жизни особей, период достижения ими половой зрелости, число генераций. Очевидно, что у многих насекомых, дающих по несколько генераций в год, возрастная структура популяции очень простая, а у долгоживущих — очень сложная.
Рождаемость в популяции определяется прежде всего эволюционным положением вида, его биологией; Низкая плодовитость характерна для тех видов, которые проявляют большую заботу ti потомстве. Кроме того, рождаемость зависит от скорости полового созревания, числа генераций в году, соотношения в популяции самцов и самок, обеспеченности кормом, влияния погодных условий.
Смертность в популяции—это количество особей, погибших за определенный период. Она бывает очень высокой и изменяется в зависимости от условий среды, возраста и состояния популяции, У большинства видов смертность в раннем возрасте всегда бывает выше, чем у взрослых особей. Однако встречаются и такие виды, у которых смертность приблизительно одинакова во всех возрастах или преобладает у особей старших возрастов. Факторы смертности очень разнообразны. Она может быть вызвана влиянием физических условий (низкие и высокие температуры, ливневые осадки и град, избыточная и недостаточная влажность и др.), биотическими факторами (отсутствие благоприятного корма, инфекционные заболевания, враги и т. д.) и антропогенными (загрязнение окружающей среды, уничтожение насекомых, вырубка деревьев и др.).
Рост популяций определяется двумя противоположными явлениями — рождаемостью и смертностью, также миграциями и иммиграциями. Большинство исследователей полагает, что рождаемость—более постоянная, хотя и очень изменчивая величина, чем' смертность. Поэтому и уровень численности зависит главным образом от факторов смертности.
Баланс популяции изучают, пользуясь таблицами выживания, которые дают возможность оценить сравнительное значение каждого фактора смертности в отдельности и выделить критические периоды в течение генерации, определяющие дальнейший ход динамики численности. В таблицах описана серия последовательных измерений, с помощью которых можно обнаружить изменение численности популяции в естественных условиях '.
Каждому виду присуща определенная оптимальная плотность популяции, отклонения от которой в обе стороны отрицательно сказываются на темпах воспроизводства и жизнедеятельности особей. Колебания численности вокруг оптимального уровня носят разный характер. Они могут быть плавными циклическими и резкими нерегулярными. Механизм этих колебаний очень сложен и до сих пор трудно прогнозируем.
• Для объяснения причин колебания численности насекомых был предложен ряд теорий, не оправдавшихся в своем первоначальном виде. Одни исследователи признавали ведущую роль в регуляции численности фитофагов за их врагами—хищниками и паразитами <паразитарная теория), другие считали, что колебания численности обусловлены воздействием на насекомых климатических факторов (климатическая теория). Ряд исследователей отдавал преимущество пищевому фактору. По мере дальнейшего развития исследований стали считать, что численность популяции каждого вида насекомого в экосистеме регулируется комплексом факторов (биоценологиче-ская теория). В настоящее время наибольшее признание получила синтетическая теория. Она рассматривает колебания численности
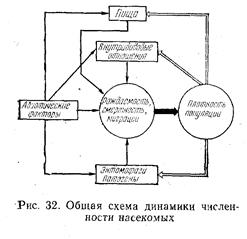 |
популяций как автоматически регулируемый процесс, слагающийся из отклонений под влиянием случайных воздействий абиотической среды и стабилизирующего действия биотических факторов. На рис. 32 показана общая схема динамики численности популяций насекомых. Существует два принципиально различных процесса, определяющих движение численности популяций во времени: модификация и регуляция. Модификация выражает подъемы и спады численности, обусловленные случайными по отношению к популяциям изменениями факторов, функционально не связанных с ее плотностью. Это в первую очередь погодные условия. Они имеют решающее значение при выяснении причин колебания численности и прогнозировании массовых размножений насекомых.
Аномальное развитие отдельных метеорологических элементов может почти мгновенно и очень резко изменить уровень численности популяции. Погодные условия влияют с равной интенсивностью при любой плотности популяции. Так, при очень низкой или очень высокой температуре, ливне или наводнении смертность будет неизбирательной и.не будет зависеть от числа особей, она, как правило,
•будет массовой, и сохранятся лишь отдельные особи в надежных укрытиях. Подъем численности популяции часто бывает обусловлен устойчивым типом погоды, например длительной засухой.
Еще чаще метеорологические условия оказывают косвенное влияние на численность популяции, усиливая смертность через другие факторы. Под влиянием погодных условий сезонные изменения развития кормовых пород могут совпадать с появлением определенных фаз данного вида насекомого или не совпадать. Так, например, в один год выход гусениц дубовой зеленой листовертки совпадает с распусканием почек дуба, а в другие годы наступает раньше, когда почки еще не раскрылись. В этом случае происходит массовая гибель гусениц. Под влиянием выпадающих осадков колеблется интенсивность смоловыделения у сосны. В годы засухи смоляное давление в хвое и почках падает, что ведет к резкому увеличению выживаемости гусениц ряда хвоегрызущих насекомых.
Регуляция численности насекомых заключается в сглаживании возникающих колебаний в результате действия регулирующих механизмов, которые зависят от плотности популяции и действуют по принципу обратной связи. Регуляторные механизмы объясняют частоту и интенсивность колебаний численности насекомых. На рис. 35 показаны основные механиз-
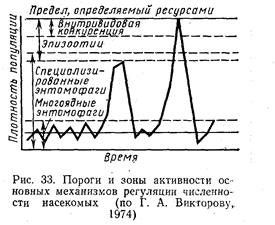 |
мы регуляции численности популяций насекомых. Различают межвидовые и внутривидовые механизмы регуляции численности. К первым относятся паразиты, хищники и патогены. Много-ядные энтомофаги (муравьи, птицы и др.) стабилизируют численность популяции на самых низких уровнях, специализированные паразиты регулируют ее на более высоких уровнях. Резкое снижение численности насекомых около ее высшего предела осуществляют болезни
насекомых, вызывающие эпизоотии. Если они отсутствуют или действуют недостаточно, включаются внутривидовые регулирующие механизмы. Они являются результатом взаимодействия особей внутри самой популяции. В простейшем виде — это разнообразные формы внутривидовой конкуренции, связанные с прямым и косвенным отрицательным воздействием особей друг на друга.
Большую роль в регуляции численности популяции играют миграции. С помощью миграций идет убыль и пополнение популяций, может резко изменяться их численность, происходить перемещение в пространстве и накопление особей в новых местах. Миграции позволяют популяции расселяться и избежать неблагоприятных условий, которые создаются в результате ее же жизнедеятельности, сохраняться в период депрессии в резервациях и затем быстро накапливаться в соседних с ними насаждениях.
Численность популяции может изменяться также и под влиянием физиологической разнокачественности особей. Например, вследствие такой разнокачественности ежегодно часть особей рыжего пилильщика, дубовой хохлатки и других вредителей леса уходит в диапаузу на фазе пронимфы и куколки, а остальные особи заканчивают нормальное развитие по одногодичному циклу. За счет этого в одних генерациях происходит быстрое сокращение, а в других — увеличение численности популяции.








