 2015-05-30
2015-05-30 2380
2380Непроизвольное внимание относится к феномену переключения внимания на стимул, который ранее не привлекал внимания. Оно протекает автоматически и не требует специальных усилий. Основу непроизвольного внимания составляет ориентировочный рефлекс (ОР). Его появление автоматически включает произвольное внимание, которое и обеспечивает дальнейшую и контролируемую обработку стимула.
Ориентировочный, или исследовательский, рефлекс был открыт И.П. Павловым и назван имрефлексом «Что такое?». Он описал его как комплекс двигательных реакций (поворот головы, глаз, настораживай ие ушей) в направлении нового стимула. Ю. Конор-
ский (1970) этому комплексу соматических реакций дал название
рефлекса прицеливания, назначение которого — обеспечить лучшее восприятие нового стимула.
Однако сначала наиболее подробно была исследована тормозная функция ОР, который всякий раз прерывал текущую деятельность собаки, подавлял ее условные рефлексы и вызывал общее торможение животного. Представление об ОР как о многокомпонентной реакции активации сложилось много позже, когда была создана и получила распространение техника полиграфической регистрации ЭЭГ, вегетативных и двигательных реакций человека и животных, хотя И.П. Павловым уже были получены первые данные о зависимости формирования условного рефлекса от наличия
ориентировочного, который обеспечивал необходимую для обучения активацию мозга.
Е.Н. Соколов (1960), а затем Р. Линн (К. Ьупп, 1966) описали систему компонентов ОР. На уровне неокортекса ОР представлен ЭЭГ-реакцией активации (блокадой альфа-ритма, усилением высокочастотных колебаний ЭЭГ), коррелирующей с ростом возбудимости, реактивности и лабильности у нейронов коры- К вегетативным компонентам ОР относят расширение зрачков, увеличение кожной проводимости, снижение ЧСС, изменение дыхания, рост дыхательной аритмии в частотном спектре ритмограммы сердца, расширение сосудов головы и сужение сосудов рук. В моторной сфере ориентировочный рефлекс, как уже отмечалось, представлен поворотом головы, движением глаз, настораживай ием ушей в направлении нового стимула и увеличением мышечного тонуса, обеспечивающего повышенную готовность к моторным реакциям. Выделен сенсорный компонент ОР, который состоит в увеличении чувствительности анализатора и может быть измерен по субъективным ответам испытуемого.
Первоначально новый стимул вызывает генерализованный ОР, связанный с возбуждением ретикулярной формации ствола мозга. Он характеризуется ЭЭГ-активацией, которая охватывает всю кору в течение достаточно длительного времени {тоническая форма ОР, часто возникающая параллельно генерализованному ОР). Генерализованный ориентировочный рефлекс быстро угасает, обычно после 10—15 применений индифферентного стимула. Это позволяет выявить локальный ориентировочный рефлекс, который более устойчив к угасанию и требует 30 и более применений стимула. Его генерацию связывают с активацией неспецифического таламуса. При локальном ОР реакция ЭЭГ-активации сохраняется в сенсорной коре, соответствующей модальности повторяемого раздражителя. Так, свет продолжает вызывать блокаду альфа-ритма, но только 62
В зрительной коре; звук — блокаду тау-ритма, регистрируемого от височной области; тактильное и проприоцептивное раздражение При пассивном подъеме пальца руки вызывает депрессию мю-рит- ма (роландического ритма) локально в сенсомоторной коре. Длительность реакции сокращается, она становится физической, появляясь только на включение и выключение стимула (фазическая реакция пробуждения). Продолжая повторять стимул, можно добиться полного угасания ОР. Ориентировочный рефлекс с полным набором компонентов возникает не у всех испытуемых, что выражает их индивидуальные различия.
Отличительной особенностью ОР является его угасание, развивающееся с повторением стимула. Однако любое изменение в повторяющемся стимуле восстанавливает ОР. Оно может быть вызвано изменением модальности, интенсивности, пространственных и временных характеристик, включая длительность стимула и меж-Стимульные интервалы. Изменения в сложных стимулах или в их комплексах, например, такие, как пропуск одного из его элементов или изменение их порядка, также восстанавливают ОР. Величина его восстановления пропорциональна числу одновременно измененных параметров и степени их изменения.
. Специфическим стимулом, на который всякий раз возникает ОР, является его абсолютная или относительная новизна. Д. Бер-лайн (О. Вег1упе) добавляет к факторам, вызывающим безусловный ОР, удивление, неопределенность, двусмысленность, сложность, необычность. Однако, по мнению Р. Линна (Ьупп К., 1966), их также можно интерпретировать как разные формы новизны. Удивление может быть вызвано появлением чего-либо нового в последовательности стимулов, а другие качества могут быть связаны с появлением новизны в паттерне стимулов.
Таким образом, ОР, обладая свойством безусловного рефлекса, возникающего на новизну, вместе с тем обнаруживает сходство и с условным рефлексом. Оно проявляется в его способности к угасанию. Угасание — это особая форма обучения, получившая название негативного научения, ведущего к потере способности данного стимула вызывать ОР.
Для объяснения механизма появления качества новизны у стимула, вызывающего ОР и последующее его угасание в результате многократного повторения стимула, Е.Н. Соколовым (1963) была предложена теория «нервной модели стимула» (рис. 14). В его концепции ключевая роль принадлежит блоку нервной модели, которая формируется в мозге только за счет повторения стимула и без специального подкрепления. В ней отражаются все параметры повторяющегося стимула (3). Нервная модель стимула многомер-
| Стимул |
| ^6 | |||||||
| Устройство, блокирующее акти ш 1ру юту ю систему | ———^ | Исполнительное устройство ориентировочною рефлекса | |||||
| Ч | 'еакци Реаш | ||||||
| 9—-. | ! Импу.| рас 001 лзс | ьсы ования | > 5 | ||||
| Вое при шее уст | нимаю- | аратор | |||||
| ройство | |||||||
| . | . 4 | ||||||
| И.. | II олн и тельное устройство условного рефлекса | ||||||
| Моделирующее устройство | <——' | ||||||
| 8 ' | |||||||
Рис. 14. Блок-схема Е.Н. Соколова, объясняющая формирование «нервной модели стимула» и появление ориентировочной реакции на «новизну». Объяснение в тексте.
на, что убедительно доказано во многих работах восстановлением ОР при изменении самых разных физических характеристик стимула. Другой ключевой структурой является наличие компаратора — устройства, выполняющего операцию сопоставления сформированной модели с сигналами сенсорной системы от предъявляемого раздражителя (2). При их несовпадении возникает сигнал рассогласования (5), активирующий исполнительное устройство ориентировочного рефлекса (7). Нервная модель стимула по мере формирования усиливает свои тормозные влияния на систему неспецифической активации в момент действия стимула, модель которого уже сформирована (6). При этом второй путь вызова ОР не работает, так как импульсы рассогласования отсутствуют. Появление ОР оказывает активирующее влияние на воспринимающее устройство (10). Предполагается, что при выработке условного рефлекса модель индифферентного стимула (будущего условного) связывается с исполнительным устройством условного рефлекса (8). Сигнал от сенсорной системы параллельно поступает к структурам, где формируется модель (3), на компаратор (2) и к неспецифической системе (1), где может развиваться привыкание — ослабление реакции активации.
Возникает вопрос: в каких структурах мозга может формироваться нервная модель стимула? Сначала Е.Н. Соколов указывал на роль коры в формировании энграмм стимулов. Однако после открытия О.С. Виноградовой (1975) в гиппокампе двух типов ней-
ронов, связанных с ОР, он стал подчеркивать роль гиппокампа в формировании нервной модели стимула. Нейроны В и Т в поле (];АЗ гиппокампа отвечают на новый стимул противоположными реакциями. Нейроны В возбуждаются, нейроны Т тормозятся. Но ®бе группы нейронов обнаруживают свойства ОР. Е.Н. Соколов предположил, что нервная модель стимула представлена параллельными матрицами потенцированных синапсов от корковых нейронов, избирательно отвечающих на физические свойства, конфигурацию сенсорного стимула, на нейронах новизны (В-нейро-ны) и тождества (Т-нейроны) гиппокампа. Реципрокные реакции нейронов новизны и тождества гиппокампа противоположно влияют на активирующую и инактивирующую системы мозга. Совместное действие обеих групп нейронов увеличивает отношение стимула к шуму, повышая тем самым надежность системы, детектирующей новизну. Реакция активации на новый раздражитель определяется возбуждением активирующей и торможением инак-тивирующей систем мозга. Привычный стимул перестает возбуждать активирующую систему и тормозить инактивирующую. Возникновение ОР на изменение привычной стимуляции объясняется тем, что оно активирует новые, еще не потенцированные (не Подвергшиеся пластическим перестройкам) синапсы нейронов новизны и тождества в гиппокампе (Соколов Е.Н., 1995).
, Процесс формирования нервной модели стимула отвечает всем Критериям, предложенным М.И. Познером и К.Р. Снайдеромдля адделения автоматически протекающих процессов. Он характеризуется непроизвольностью и протекает без усилий, не осознается и не влияет на текущую деятельность. В процессе формирования модели информация обо всех физических свойствах стимула обрабатывается параллельно, без ограничений их количества. Это Происходит и тогда, когда внимание субъекта отвлечено от стимула и направлено на какую-то деятельность. Операции сличения действующего раздражителя с моделями памяти также протекают автоматически. Таким образом, безусловный ОР — конечное звено серии операций, представляющих автоматическую обработку информации.
Концепция Е.Н, Соколова о нервной модели стимула и экспериментальные данные, подтверждающие ее, хорошо согласуются с теориями внимания — ранней и поздней селекции. Обе теории связывают начальный этап обработки сенсорной информации с параллельно и автоматически протекающими процессами. Их различает лишь представление об обработке семантической информации. По теории ранней селекции (фильтрации) семантическая информация требует произвольного внимания, в то время как тео-
5-3341
рия поздней селекции допускает, что часть значимой семантической информации может обрабатываться автоматически.
Результаты опытов Е.Н. Соколова с изучением семантического поля ориентировочного рефлекса показывают, что анализ семантических связей также может осуществляться автоматически, на неосознаваемом уровне, и вызывать ОР, когда предъявляется слово, близкое по звучанию к угашенному, но сильно отличающееся от него по смыслу (Данилова Н.Н., Крылова А.Л., 1997).
Иное объяснение угасания ОР предложено Р. Томпсоном и др. (Тпотрзоп К. е1 а1., 1973) в его двухпроцессной модели. В отличие от модели Е.Н. Соколова его модель построена на результатах изучения механизма привыкания безусловных рефлексов. Для этого в качестве объекта исследования он выбрал двигательную реакцию — рефлекс сгибания задней лапы у спинальной кошки в «остром эксперименте» (препарат с рассеченным спинным мозгом). Было показано, что рефлекс при многократном повторном вызове обнаруживает привыкание со всеми свойствами, характерными для угасания ОР. И этот процесс не требует участия головного мозга. С точки зрения Р. Томпсона, привыкание и растор-маживание можно объяснить, не прибегая к концептам модели и компаратора. Он предположил, что величина реакции зависит от соотношения двух процессов: привыкания и сенситизации- Привыкание развивается в результате повторения стимула и ведет к уменьшению ответа, а сенситизация — к его увеличению. Сенси-тизация зависит от состояния организма, т.е. от уровня его активации. Оба процесса предполагаются независимыми (рис. 15). После первых стимулов сенситизация растет, затем постепенно падает, так как и у нее развивается привыкание. Второй процесс — привыкание — развивается более монотонно. Восстановление реакции на изменение стимула объясняют тем, что привыкание (ослабление) сенситизации еще не стало генерализованным, т.е. стимулы, сильно отличающиеся от повторяющегося, способны вызывать эффект сенситизации. Чем меньше общих путей проведения возбуждения для нового и старого стимулов, тем сильнее ответ на изменение.
Модель Р. Томпсона относится к одностадийным, так как привыкание реакции возникает на путях между стимулом и реакцией. Теория Е.Н. Соколова двухстадийна, она не только рассматривает пути между стимулом и реакцией, но и вводит дополнительные структуры (модель, компаратор). Уязвимым местом модели Р. Томпсона является то, что она была построена на данных о привыкании только одной реакции — двигательного рефлекса сгибания лапы. Кроме того, привыкание, исследованное Р. Томпсо-66
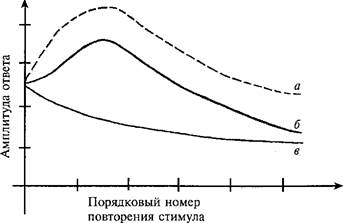
Рис. 15. Зависимость амплитуды сгибательного рефлекса от баланса процессов сенситизации и привыкания в ситуации повторения стимула I (по К.. ТЬотрзоп е1 а1., 1973).
а — сенситизация; б ~ экспериментальные данные; в — привыкание.
ном у декортицированного животного, развивалось крайне медленно по сравнению с быстрым привыканием компонентов ОР. Скорее всего, это два разных механизма, которые существуют параллельно. Быстрое привыкание (угасание ОР) требует сохранности головного мозга, медленное — может происходить у нейронов спинного мозга.
В теории А.С. Бернстайна (Ветп51ет А.5., 1969) о значимости стимула как главного фактора, определяющего появление ОР, модель Е.Н. Соколова дополняется еще одной стадией обработки Информации, которая следует за операцией, выполняемой компаратором (трехстадийная модель). Это — стадия оценки «значимости^ стимула. Факт, что восстановление ОР после его угасания происходит несимметрично на увеличение и уменьшение интенсивности стимула (более сильный раздражитель вызывает больший ориентировочный рефлекс), объясняется тем, что новый стимул большей интенсивности может указывать на опасность и поэтому является более значимым по сравнению с новым, но слабым стимулом. А. Бернстайн утверждает, что ОР — не столько реакция на новизну, сколько результат принятия решения о том, что полученная информация, возможно, будет полезной для субъекта.
Однако в модели Е.Н. Соколова значимость также является характеристикой, вызывающей ОР, и она представлена условным
5*
ориентировочным рефлексом, Он показал, что придание раздражителю сигнального значения обычно с помощью инструкции или речевого подкрепления, требующих отвечать определенной реакцией на определенный стимул, восстанавливает или усиливает ОР на условный сигнал. Совпадение условного стимула с его моделью в этом случае вызывает не подавление ОР, а, наоборот, его усиление. И этот эффект сохраняется, пока условный рефлекс не достигнет стадии автоматизации.
5.5. ПОТЕНЦИАЛЫ, СВЯЗАННЫЕ С СОБЫТИЯМИ,

