2015-06-10
2015-06-10 705
705Спектрально-корреляционный анализ электрической активности различных образований головного мозга значительно пополнил сведения о функциональной организации ряда форм высшей нервной деятельности животных и человека. Наиболее обстоятельно были изучены пространственные отношения процессов, протекающих в различных мозговых структурах при ориентировочной реакции, при выработке и угашении условных рефлексов, в естественном и медикаментозном сне, при некоторых умственных операциях, совершаемых здоровым и больным человеком [Ливанов, 1972]. Значительно меньше известно о пространственной организации биоэлектрических процессов при возникновении состояний голода, жажды, полового влечения и во время поведения, направленного на удовлетворение этих биологических потребностей.
Хорошей моделью для изучения мотивированного поведения и его электрофизиологических коррелятов является феномен самораздражения. Однако традиционный способ самостимуляции ритмическим током исключает возможность изучения электрической активности раздражаемых структур, хотя возбуждение именно этих структур инициирует все последующие действия животного. Вот почему использование постоянного тока для самостимуляции животных впервые открыло принципиально новые возможности электрофизиологического исследования этой формы мотивированного поведения [Павлыгина, Михайлова, Симонов, 1975].
Систематическими работами В. С. Русинова [1969] и его сотрудников были показаны преимущества постоянного тока для создания стойких доминантных очагов в центральной нервной системе. Поляризуя гипоталамическую область головного мозга кролика, Р. А. Павлыгина [1956, 1958] получила доминантный очаг, оказывающий выраженное влияние на оборонительные условные рефлексы, электрическую активность различных мозговых образований и кровяное давление. Приступая к экспериментам, мы не нашли в литературе работ, посвященных поляризации эмоциональных «центров» и феномену самораздражения постоянным током, равно как и данных о пространственной организации электрической активности определенных образований мозга при мотивированном поведении животных. Совместно с Р. А. Павлыгиной, В. Д. Трушом и Н. Г. Михайловой опыты были поставлены нами на 16 белых крысах-самцах. У 6 из них на 6-ти канальном магнитофоне производилась регистрация суммарной электрической активности моторной и зрительной областей коры, гиппокампа, обонятельных луковиц, эмоционально-позитивных и негативных зон гипоталамуса (крысы № 13, 14, 15, 197, 221, 222).
Для монополярного раздражения использовали электроды с диаметром кончика 60 мкм, зачищенным от лака на 0,3-0,5 мм. Индифферентный электрод находился в мышцах шеи. Для получения феномена самостимуляции раздражали структуры медиального пучка переднего мозга в преоптической области и латеральном гипоталамусе. Эмоционально негативные реакции получали при раздражении дорзомедиальной области покрышки. Прежде чем поляризовать позитивные зоны, эти точки тестировались на эффективность в отношении самораздражения ритмическим током. С этой целью использовали однофазные, прямоугольные электрические импульсы частотой 1000 циклов в секунду при длительности стимулов 0,1 мс, длительности пачки 0,3- 0,5 с интенсивностью 240-560 мкА. Поляризация малыми токами 2-6 мкА, которая обычно используется для создания доминантных очагов при экспозиции около одного часа, была неприемлема в наших экспериментах. Мы использовали токи 20-100 мкА и животное не находилось на педали дольше 3 мин.
Спустя 10 дней после операции оживления электродов крысу помещали в плексиглазовый ящик размерами 21X25X40 см, у одной из стенок которого находилась педаль. Как правило, на протяжении первых 2-3 дней регистрировали количество нажатий на педаль, совершаемых крысой в процессе ее ориентировочно-исследовательской активности. Затем замыкали цепь, в результате чего каждое нажатие на педаль начинало сопровождаться воздействием анода или катода постоянного тока. Поляризации подвергали только те пункты, подкрепляющие свойства которых были предварительно установлены стимуляцией ритмическим током. Если крыса долго не вставала на педаль, ее сажали на педаль, удерживая в этом положении несколько секунд. Об эффективности поляризации эмоционально-позитивных зон гипоталамуса можно было судить по числу нажатий и продолжительности пребывания на педали, а также - по динамике угашения реакции самостимуляции после выключения тока. В ряде случаев влияние постоянного тока контролировалось по избеганию нажатий на педаль после поляризации эмоционально-негативных пунктов «наказания».
Четырехсекундные интервалы записи электрической активности мозговых образований обрабатывали на ЭЦВМ. При этом оценивали автоспектры потенциалов каждого исследуемого образования, а также кросскорреляционные коэффициенты и когерентные функции между потенциалами каждой пары изучаемых структур. Все спектральные характеристики оценивали в диапазоне от 1 до 20 Гц. Методика определения частотного спектра и вычисления функции когерентности описаны ранее [Труш, Кориневский, 1978]. Морфологический контроль мозга крыс подтвердил локализацию отводящих и стимулирующих электродов.
У крыс № 197, 221 и 222, у которых регистрировалась электрическая активность двигательной и зрительной областей новой коры, эмоционально-позитивной и негативной зон гипоталамуса, в фоновых потенциалах перечисленных структур согласно автоспектральным оценкам, как правило (примерно 80% наблюдений), доминирующими были частоты тета- и альфа-диапазонов. Во время пребывания крысы на педали при действии подкрепляющего постоянного тока в 40% записей наблюдалось доминирование дельта-волн. Приблизительно в 50% случаев, когда наиболее выраженными оставались колебания тета- и альфа-диапазонов, частота доминирующего ритма уменьшалась на 1,5-3 Гц по сравнению с фоновыми записями. После ухода крысы с педали во всех без исключения случаях частота доминирующего ритма возрастала в 60% -на 0,5-2 Гц.
При анализе оценок когерентных функций гипоталамических и корковых структур прежде всего обращает на себя внимание то, что каждому из изучавшихся в эксперименте этапов поведения соответствует определенный «рисунок» распределения корреляционных связей (рис. 36). Это подтверждает мнение М. Н. Ливанова [1972] о том, что именно в пространственной организации биопотенциалов находят отражение различные функциональные состояния мозга, что было убедительно показано, например, для ориентировочного рефлекса, для различных стадий выработки условного рефлекса и для некоторых тормозных состояний.
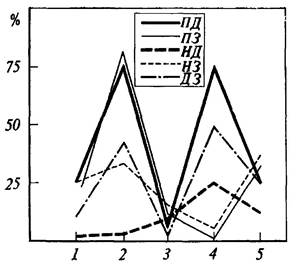
Рис. 36. Оценки вероятности наличия значимой когерентности между потенциа лами мозговых структур на различных этапах поведения при осуществлении реакции самополяризации
Ордината - процент случаев регистрации значимой когерентности на тета- и альфа-частотах; абсцисса - этапы поведения крысы: 1-фон, 2-перед нажатием на педаль; 3 -во время пребывания на педали, 4 -перед уходом с педали, 5 - после ухода с педали. Кривыми обозначено поведение когерентности для потенциалов различных структур; П - эмоционально-позитивный пункт гипоталамуса; Я - эмоционально-негативный пункт; Д и 3 - соответственно двигательная и зрительная области коры (по Р. А. Павлыгиной, В. Д. Трушу, Н. Г. Михайловой и П. В. Симонову).
В табл. 5, где представлены результаты анализа оценок когерентных функций потенциалов мозга крыс № 221 и 222, приняты следующие обозначения. Цифры показывают процент случаев регистрации значимой когерентности (уровень значимости Р<0,05) на частотах тета- и альфа-диапазонов при наличии соответствующих ритмических составляющих в потенциалах сравниваемых образований.
Таблица 5
Наличие значимой когерентности между потенциалами гипоталамических и корковых структур мозга на различных этапах поведения при осуществлении и угашении реакции самораздражения, %
Примечание. П - эмоционально-позитивный пункт гипоталамуса, Н - эмоционально-негативный пункт, Д, 3 - соответственно двигательная и зрительная область коры.
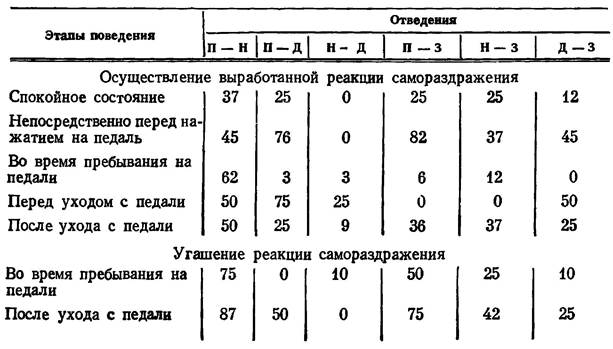
Рассматривая таблицу, можно убедиться, что непосредственно перед нажатием на педаль резко (более чем в три раза) возрастает когерентность электрической активности эмоционально-позитивного пункта гипоталамуса с ЭЭГ моторной и зрительной коры и когерентность потенциалов двух корковых участков. В то же время корреляционные отношения между потенциалами этих структур и эмоционально-негативной зоны гипоталамуса существенно не изменяются.
Напомним, что согласно М. Н. Ливанову [1972, с. 169] «...в синхронности колебаний биопотенциалов каких-либо пунктов коры головного мозга мы видим не прямое выражение связей между ними, а лишь условия, делающие возможной их реализацию». Возросшая в нашем случае когерентность, по-видимому, свидетельствует о готовности проведения возбуждения по двум каналам: от ранее раздражавшегося эмоционально-позитивного пункта к двигательной коре и к зрительному анализатору, воспринимающему условный сигнал будущего подкрепления (вид педали, ее местонахождение в камере и т. д.). Более чем в три раза возрастает также когерентность ЭЭГ двигательной и зрительной коры, поскольку именно вид педали направляет движение животного, инициированное следовым возбуждением эмоционально-позитивной зоны.
В период пребывания крысы на педали когерентность между потенциалами практически всех изучавшихся структур падает: животное получает подкрепление постоянным током и остается совершенно пассивным. Такое падение когерентности наряду с наблюдающимися на этом этапе замедлением ритмики потенциалов и усилением дельта-активности делают состояние животного во время самостимуляции постоянным током весьма сходным по электрографическим показателям с такими тормозными состояниями, как естественный, наркотический и электросон. Для ситуации самостимуляции характерным при этом является возрастание когерентностиэмоционально-позитивных и негативных структур, что, возможно, является отражением постепенной замены эмоционально-позитивного состояния эмоционально-негативным возбуждением, побуждающим животное прервать стимуляцию и уйти с педали.
Непосредственно перед уходом крысы с педали впервые возрастает когерентность потенциалов негативного пункта и моторной области. Одновременно увеличивается когерентность потенциалов моторной и зрительной коры, а также - моторной коры и позитивного пункта наряду с нулевой когерентностью электрической активности зрительной коры и гипоталамуса. В этой картине пространственной организации биоэлектрической активности находят отражение некоторые черты формирования новой программы действий. Причем, характерным отличием этой новой двигательной задачи - реакции избегания - от задачи, формировавшейся перед нажатием на педаль, являются, прежде всего, иная конечная цель и возбуждение иного мотивационного центра, инициирующего двигательную активность. Не исключено, что в зарегистрированной на данном этапе совокупности электрографических характеристик отражается и такая особенность состояния мозга, как своеобразная борьба двух тенденций: остаться на педали ради продолжения подкрепления или уйти с нее, поскольку действие постоянного тока становится все более аверсив-ным. С этим могут быть связаны, в частности, активация не только негативного пункта гипоталамуса (в приблизительно 30%
наблюдений - учащение ритмики потенциалов и ни одного случая урежения) и увеличение корреляционной зависимости между электрической активностью негативной и позитивной зон гипоталамуса, с одной стороны, и ЭЭГ моторной коры - с другой. После ухода крысы с педали когерентность потенциалов практически всех исследованных структур возвращается к уровню, характерному для ситуации, предшествовавшей началу выработки реакции самораздражения. При этом несколько увеличенная по сравнению с фоном когерентность потенциалов негативного пункта и двигательной области коры возможно связана с наличием следов эмоционально-негативного состояния.
Весьма характерная картина наблюдается на первых этапах угашения, когда крыса, надавив на педаль, не получает подкрепления постоянным током. Здесь возрастает когерентность потенциалов позитивного пункта и зрительной коры, свидетельствуя о необходимости продолжения поиска отсутствующего подкрепления. После ухода крысы с педали мозг ее остается в состоянии повышенной готовности к новым действиям: пространственная организация биоэлектрической активности практически идентична той, что наблюдалась в ситуации перед началом движения животного к педали. Максимальных величин достигает возросшая еще во время пребывания крысы на педали когерентность позитивного и негативного пунктов, отражая «борьбу» между состоянием неудовлетворенности и стремлением к будущему подкреплению.
До сих пор мы анализировали данные, касающиеся электрической активности эмоционально-позитивных и негативных зон гипоталамуса, а также моторной и зрительной областей коры. Каково участие в осуществлении мотивированного поведения структур гиппокампа и обонятельной луковицы, которые у крыс принадлежат к мозговому субстрату поисковых действий, не обязательно связанных с анализом обонятельных стимулов? Соответствующий фактический материал был получен при анализе электрической активности, отводимой одновременно от гиппокампа, эмоционально-позитивного пункта гипоталамуса, обонятельной луковицы и зрительной области коры у крыс № 13, 14 и 15.
Изменения оценок автоспектральных функций потенциалов исследуемых образований были на всех этапах в значительной мере аналогичны тем, что описаны ранее для электрической активности гипоталамических и корковых структур у крыс № 197, 221 и 222. Некоторые отличия отмечены лишь для потенциалов обонятельной луковицы, в которых практически всегда доминировали дельта-волны. Однако и здесь можно было заметить некоторое учащение и замедление доминирующей ритмики параллельно с соответствующими изменениями в потенциалах гиппокампа, гипоталамуса и коры.
Результаты анализа оценок когерентных функций приведены в табл. 6 на примере данных, полученных в экспериментах с кры сой № 13. На крысах № 14 и 15 получены аналогичные результаты. Табл. 6 построена по тому же принципу, что и табл. 1.
Таблица 6
Наличие значимой когерентности между потенциалами гиппокампа, гипоталамуса, обонятельной луковицы и коры на различных этапах поведения при осуществлении и угашении реакциии самораздражения
Примечание. Г - гиппокамп, П - эмоционально-позитивный пункт гипоталамуса, Б - обонятельная луковица, 3 - зрительная область коры.
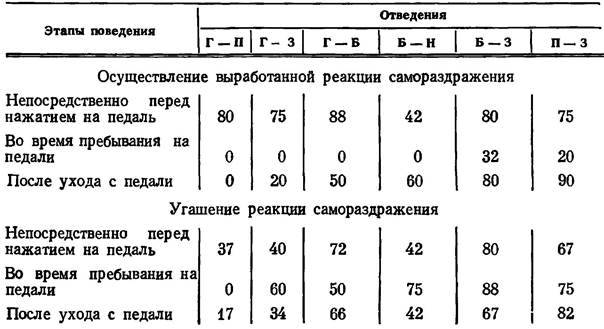
Сопоставляя распределение корреляционных отношений между потенциалами изучаемых структур при осуществлении реакции самораздражения и в процессе ее угашения, мы обнаруживаем наибольшее различие в период, когда крыса находится на педали. Во время подкрепления когерентность падает между потенциалами всех образований. Если же крыса, находясь на педали, не получает подкрепления постоянным током, когерентность возрастает, особенно между потенциалами позитивного пункта и обонятельной луковицы (почти в два раза), гиппокампа и зрительной коры (на 20%). При анализе этих данных следует учитывать, что наблюдаемая организация электрической активности должна отражать, в частности, такую особенность состояния ЦНС, как поиск неудовлетворенным животным отсутствующего подкрепления. После ухода крысы с педали организуется новая система корреляционных отношений электрической активности изучаемых образований мозга. Характерным при этом является то обстоятельство, что синхронность потенциалов гиппокампа с потенциалами всех исследуемых структур оказывается более высокой, чем это наблюдается у животного, получившего подкрепление постоянным током. Гиппокамп, таким образом, в этой ситуации становится одним из центральных образований в системе структур, электрическая активность которых связана значимой
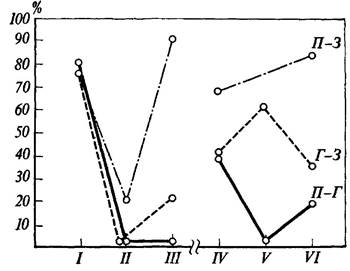
Рис. 37. Вероятность наличия значимой когерентности между потенциалами мозговых структур на различных этапах поведения при осуществлении реакции самополяризации
корреляционной зависимостью. Изменения процента случаев значимой когерентности на тета- и альфа-частотах у крысы № 13 приводятся на рис. 37.
Ордината -• процент случаев регистрации значимой когерентности в тета- и альфа-частотах; абсцисса - этапы поведения крысы: I - перед нажатием на педаль, II - во время пребывания на педали, III - после ухода с педали, IV - перед нажатием на педаль в опыте с угашением, V - во время пребывания на педали в опыте с угашением, VI - после ухода с педали. Кривыми обозначено изменение когерентности для потенциалов различных структур: П - эмоционально-позитивный пункт гипоталамуса, З - зрительная область коры, Г - гиппокамп (по Р. А. Павлыги-ной, В. Д. Трушу, Н. Г. Михайловой и П. В. Симонову).
Результаты опыта удобнее рассмотреть, пользуясь схематическим рис. 38. На этой схеме сплошными линиями обозначен высокий (свыше 50) процент случаев значимой когерентности, а прерывистыми линиями средний (от 20 до 50) процент. Низкий (меньше 20) процент линиями не обозначен. Если принять основной постулат концепции М. Н. Ливанова о том, что «...в формирующихся по мере становления условной связи корково-подкорковых взаимоотношениях мы видим не пути замыкания рефлекса, но пути тех влияний, которые определяют возможность передачи возбуждения в коре головного мозга» [Ливанов, 1972, с. 88], то динамика функциональных взаимоотношений трех исследованных структур окажется весьма интересной.
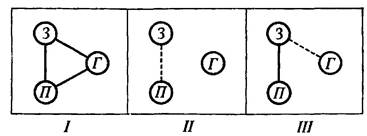
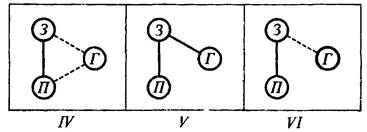
Рис. 38. Схематическое изображение взаимоотношений электрической активности мозговых структур
Сплошные линии - процент случаев значимой когерентности больше 50%; прерывистые линии - от 50 до 20%; отсутствие линий - меньше 20%. Остальные обозначения те же, что на рис. 37.
Мы видим, что на всех этапах поведения наиболее стабиль а связь эмоционально-позитивных структур гипоталамуса со зрительной корой больших полушарий головного мозга. В этой функциональной зависимости мы усматриваем проявление двусторонней условной связи между структурами, воспринимающими условный сигнал, и структурами, к которым адресуется подкрепляющее воздействие. Важнейшее значение такого рода условных связей в рефлекторных механизмах мотивированного поведения интенсивно изучается Э. А. Асратяном. Достаточно стабильно и взаимодействие между зрительной корой и гиппокампом. Наиболее интересна функциональная зависимость между гипоталамусом и гиппокампом: высокий процент случаев значимой когерентности здесь регистрируется только на одном этапе поведения-непосредственно перед тем, как крыса сядет на педаль, замыкающую ток. На всех остальных этапах этот процент ниже 20.
Мы не вырабатывали условного двигательного рефлекса самораздражения у гиппокампэктомированных крыс. Что касается естественных пищевых и оборонительных условных рефлексов, то, как мы убедились в этом выше, они вырабатываются после разрушения гиппокампа достаточно хорошо. Опыт с самополяризацией важен для нас пока только в одном отношении: он еще раз демонстрирует наличие поведенчески значимого взаимодействия между мотивационно-эмоциональными структурами гипоталамуса и гиппокампом. Анатомические связи между этими образованиями прослежены достаточно хорошо: через задний и переднелатеральный гипоталамус идет основной восходящий путь активации гиппокампа [Виноградова, 1975, с. 171].