 2015-06-05
2015-06-05 898
898Память человека часто является ассоциативной; один предмет напоминает нам о другом, а этот другой о третьем. Если позволить нашим мыслям, они будут перемещаться от предмета к предмету по цепочке умственных ассоциаций. Кроме того, возможно использование способности к ассоциациям для восстановления забытых образов. Если мы забыли, где оставили свои очки, то пытаемся вспомнить, где видели их в последний раз, с кем разговаривали и что делали. Посредством этого устанавливается конец цепочки ассоциаций, что позволяет нашей памяти соединять ассоциации для получения требуемого образа.
Ассоциативная память Хопфилда является, строго говоря, автоассоциативной, это означает, что образ может быть завершен или исправлен, но не может быть ассоциирован с другим образом. Данный факт является результатом одноуровневой структуры ассоциативной памяти, в которой вектор появляется на выходе тех же нейронов, на которые поступает входной вектор.
Двунаправленная ассоциативная память (ДАП) является гетероассоциативной; входной вектор поступает на один набор нейронов, а соответствующий выходной вектор вырабатывается на другом наборе нейронов. Как и сеть Хопфилда, ДАП способна к обобщению, вырабатывая правильные реакции, несмотря на искаженные входы. Кроме того, могут быть реализованы адаптивные версии ДАП, выделяющие эталонный образ из зашумленных экземпляров. Эти возможности сильно напоминают процесс мышления человека и позволяют искусственным нейронным сетям сделать шаг в направлении моделирования мозга.
Структура ДАП
На рис. 27 приведена базовая конфигурация ДАП. Она выбрана таким образом, чтобы подчеркнуть сходство с сетями Хопфилда и предусмотреть увеличение количества слоев.
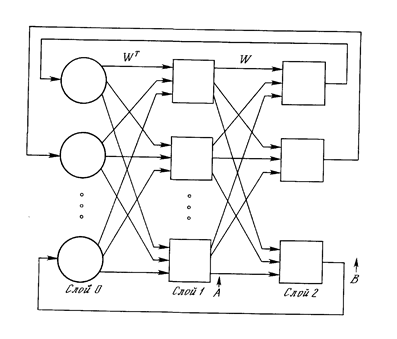
Рис. 27. Конфигурация двунаправленной ассоциативной памяти.
Входной вектор А обрабатывается матрицей весов W сети, в результате чего вырабатывается вектор выходных сигналов нейронов В. Вектор В затем обрабатывается транспонированной матрицей Wt весов сети, которая вырабатывает новые выходные сигналы, представляющие собой новый входной вектор А. Этот процесс повторяется до тех пор, пока сеть не достигнет стабильного состояния, в котором ни вектор А, ни вектор В не изменяются. Заметим, что нейроны в слоях 1 и 2 функционируют, как и в других парадигмах, вычисляя сумму взвешенных входов и вычисляя по ней значение функции активации F. Этот процесс может быть выражен следующим образом:
B=F(AW),
где В - вектор выходных сигналов нейронов слоя 2,
А -вектор выходных сигналов нейронов слоя 1,
W - матрица весов связей между слоями 1 и 2,
F - функция активации.
Аналогично
A = F(BWt),
где Wt является транспозицией матрицы W.
В качестве активационной функции в простейшем случае используется пороговая функция.
Примем также, что внутри каждого нейрона существует память в слоях 1 и 2 и что выходные сигналы нейронов изменяются одновременно с каждым тактом синхронизации, оставаясь постоянными между этими тактами. Таким образом, поведение нейронов может быть описано следующими правилами:
OUTi(n+l)=l, если NETi (n) > О,
OUTi(n+l)=0, если NETi(n)<0,
OUTi(n + 1) = OUT(n), если NETi(n) = О,
где OUTi(n) представляет собой величину выходного сигнала нейрона i в момент времени n.
Заметим, что в описанных ранее сетях слой 0 не производит вычислений и не имеет памяти; он является только средством распределения выходных сигналов слоя 2 к элементам матрицы Wt.








