2015-06-26
2015-06-26 452
4521.
А) Влияние законы действия масс. В катализируемой ферментом обратимой химической реакции, например А + В <=> С + D, концентрация компонентов реакции и соответственно направление реакции будут регулироваться влиянием закона действия масс. Оно, в частности, может быть показано в обратимой реакции трансаминирования, катализируемой ферментом аланинаминотрансферазой: Аланин + α-Кетоглутарат <=> Пируват + Глутамат. Б) Изменение количества фермента. Замена в среде глюкозы на лактозу (индуктор) приводит к индуцированному или адаптивному (после небольшого периода лаг-фазы) синтезу фермента галактозидазы (програм- мированному лактозным геном, см. главу 13), расщепляющей лактозу на глюкозу и галактозу. В клетках прокариот и эукариот имеются ферменты, концентрация которых не требует добавления индуктора; это так называемые конститутивные ферменты. Количество фермента в клетке зависит от наличия продукта реакции, катализируемой данным ферментом, причем продукт реакции вызывает торможение синтеза фермента в результате репрессии. В) Проферменты. Протеолитические ферменты пищеварительного тракта, а также поджелудочной железы синтезируются в неактивной форме – в виде проферментов (зимогенов). Регуляция в этих случаях сводится к превращению проферментов в активные ферменты под влиянием специфи- ческих агентов или других ферментов – протеиназ. Примерами подобного активирования белков является активирование некоторых гормонов (проинсулин —> инсулин), белка соединительной ткани (растворимый проколлаген превращается в нерастворимый колла- ген), белков свертывающей системы крови. Г) Химическая модификация фермента. различают обратимую ковалентную и нековалентную химические модификации ферментов, осуществляемые через ОН-группы серина, реже – тирозина или за счет нековалентных взаимодействий с молекулой фермента. В первом случае активным ферментом оказывается или фосфорилированная, или дефосфорилированная форма, как в случае с молекулами мышечной фосфорилазы и гликогенсинтазы соответственно. Д) Аллостерическая регуляция. Во многих строго биосинтетических реакциях основным типом регуляции скорости многоступенчатого ферментативного процесса является ингибирование по принципу обратной связи. Это означает, что конечный продукт биосинтетической цепи подавляет активность фермента, катализирующего первую стадию синтеза, которая является ключевой для данной цепи реакции. Поскольку конечный продукт структурно отличается от субстрата, он связывается с аллостерическим (некаталитическим) центром молекулы фермента, вызывая ингибирование всей цепи синтетической реакции.
2.
Название ферментов составляют из химического названия субстрата и названия той реакции, которая осуществляется ферментом. Если химическая реакция, ускоряемая ферментом, сопровождается переносом группировки атомов от субстрата к акцептору, название фермента включает также химическое наименование акцептора. Например, пиридоксальфермент, катализируюший реакцию переаминирования между L-аланином и альфа-кетоглутаровой кислотой, называется L-аланин: 2-оксоглутарат аминотрансфераза. В этом названии отмечены сразу три особенности: 1) субстратом является L-аланин; 2) акцептором служит 2-окcоглутаровая кислота; З) от субстрата к акцептору передается аминогруппа.
Каждому ферменту присвоен индивидуальный номер (шифр). Например, шифр уреазы выражается цифрами 3.5.1.5. Это означает, что уреаза относится к 3-му классу (первая цифра) ферментов, все представители которого катализируют реакции гидролиза. Вторая цифра (5) говорит о том, что уреаза принадлежит к 5-му подклассу этого класса, куда зачислены все ферменты, ускоряющие гидролиз С - N-связей, не являющихся пептидными. Третья цифра шифра (1) указывает на принадлежность уреазы к подподклассу 5-го подкласса, члены которого ускоряют гидролиз линейных амидов, а последняя цифра (5) - порядковый номер уреазы в этом подподклассе.
3.
Оксидоредуктазы. К классу оксидоредуктаз относят ферменты, катализирующие с участием двух субстратов окислительно-восстановительные реакции, лежащие в основе биологического окисления. Систематические названия их составляют по форме ≪донор: акцептор оксидоредуктаза≫. Например, лактат: НАД+ оксидоредуктаза для лактатдегидрогеназы (ЛДГ). Различают следующие основные оксидоредуктазы: аэробные дегидрогеназы или оксидазы, катализирующие перенос протонов (электронов) непосредственно на кислород; анаэробные дегидрогеназы, ускоряющие перенос протонов (электронов) на промежуточный субстрат, но не на кислород; цитохромы, катализирующие перенос только электронов. К этому классу относят также гемсодержащие ферменты каталазу и пероксидазу,катализирующие реакции с участием перекиси водорода.
4.
Определение активности амилазы в биологических жидкостях (моча, сыворотка крови, слюна) основано на нахождении наименьшего количества фермента, необходимого для полного расщепления добавленного крахмала в стандартных условиях. Активность фермента измеряется амилазными единицами, т. е. числом миллилитров 0,1% раствора крахмала, расщепленного в течение 30 мин. при t° 45 9 1 мл исследуемого раствора. Например, в норме активность амилазы в моче равна 16—64. Повышенные значения наблюдаются при панкреатите, заболеваниях желчных путей и др., пониженные значения вплоть до нуля — при почечной недостаточности.
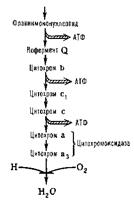
К числу ферментов дыхательной цепи относятся: 1. Пиридинзависимые дегидрогеназы с коферментом НАД + и. НАДФ+; 2. Флавинзависимые дегидрогеназы (флавопротеиды), у которых простетической группой служат ФМН и ФАД; 3. Цитохромы и цитохромоксидаза, их простетической группой является гем.