 2015-06-28
2015-06-28 1678
1678| Пространственная конфигурация молекул ДНК была установлена в 1953 г. Это открытие было удостоено высшей научной награды — Нобелевской премии. Согласно модели, предложенной Дж.Уотсоном и Ф. Криком, молекула ДНК представляет собой две параллельные неразветвленные полинуклеотиднъте цепи, закрученные вокруг общей оси в двойную правостороннюю спираль. Диаметр двойной спирали равен 20 нм с шагом около 3,4 нм, на каждый виток приходится 10 пар нуклеотидов, поскольку расстояние между соседними парами составляет 0,34 нм (рис. IV.6). Направление цепей взаимно противоположно. На 5'-конце цепи ДНК всегда расположен свободный фосфат у 5'-атома углерода; на противоположном 3'-конце — свободная ОН-группа у З'-атома углерода. Сахарно-фосфатный остов располагается по периферии спирали, а азотистые основания находятся внутри ее. Между основаниями образуются специфические водородные связи (рис. IV.7). В 50-е годы было установлено, что в ДНК, выделенной из клеток различных организмов, молярная сумма пуриновых оснований (аденин + гуанин) всегда равна сумме пиримидиновьгх (цитозин + тимин). Содержание аденина равно содержанию тимина, а содержание гуанина — количеству цитозина (правило Э.Чаргаффа). | 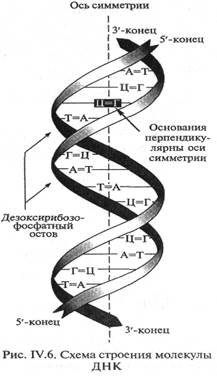 | |
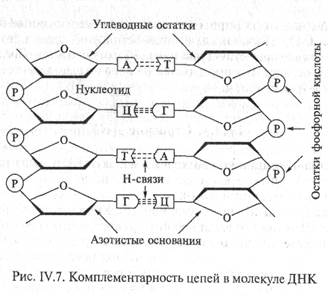 | 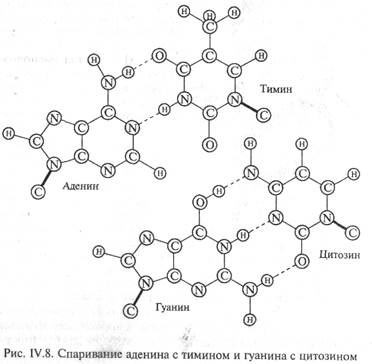 | |
Водородные связи возникают между пуриновым основанием одной цепи и пиримидиновым основанием другой цепи в результате избирательного спаривания оснований. Аденин всегда образует водородные связи с тимином (А-Т), а гуанин с цитозином (Г-Ц). Правило образования двунитевой структуры ДНК (А-Т, Г-Ц) называется правилом комплементарности (лат. complementum — дополнение). Образование водородных связей между комплементарными парами обусловлено их пространственным соответствием (рис. IV.8). Это приводит к тому, что нуклеотидные последовательности двух антилараллельных цепей ДНК всегда строго комплементарны друг к другу, а порядок чередования нуклеотидов в обеих цепях ДНК оказывается взаимообусловленным. Именно комплементарностью определяется точное воспроизведение последовательности оснований при копировании (репликации) молекул ДНК.
Водородные связи между парами комплементарных нуклеотидов (две для пары А-Т и три для пары Г-Ц) относительно непрочные. Поэтому комплементарные нити молекулы ДНК могут разделяться и соединяться вновь при изменении некоторых условий (например, изменении температуры или концентрации солей). Разделение двухцепочечной ДНК называется денатурацией, а обратный процесс - образование двухцепочечной структуры ДНК - гибридизацией Очевидно, что в реакцию гибридизации могут вступать только комплементарные друг другу нуклеотидные последовательности одноцепочечных молекул.
Цепь, содержащая информацию о строении белка (в направлении 5'-3'), называется смысловой цепью, а комплементарная - антисмысловой. Антисмысловая цепь имеет большое значение при стабилизации структуры двойной спирали ДНК и участвует в процессах репликации и репарации (восстановления) поврежденных участков ДНК. Молекулы ДНК являются гигантскими полимерами. Единицами измерения длины молекулы приняты: пары нуклеотидов (п.н.), тысячи пар нуклеотидов - килобазы (кб), миллионы пар оснований - мегабазы (мб).
У человека гаплоидный набор содержит 3,2х109 пар нуклеотидов, или 3,2 миллиарда пар оснований. Почти вся ДНК клетки содержится в ядре в виде 46 плотно упакованных, суперскрученных за счет взаимодействий с ядерными белками, структурах - хромосомах. Сравнительно небольшая часть ДНК (около 5%) локализована в митохондриях.








