 2015-06-26
2015-06-26 4673
4673В 1961 г. М. Мезельсон и Дж. Вейгл на примере фага λ показали, что рекомбинация (кроссинговер) включает разрыв и последующее воссоединение молекул ДНК. Эта работа дала импульс поиску ферментов, участвующих в процессе рекомбинации. Вскоре при изучении фагов λ и Т4 были выявлены фагоспецифичные нуклеазы, необходимые для осуществления фаговой рекомбинации. Это указывало на правильность предложенной гипотезы о механизме кроссинтовера. Начались интенсивные поиски фермента, участвующего в воссоединении расщепленных нуклеазами молекул ДНК. В 1967 г. независимо в нескольких лабораториях был открыт фермент, названный ДНК-лигазой, который катализирует синтез фосфодиэфирной связи в двухцепочечной ДНК.
Удалось обнаружить два типа ДНК-лигаз: фермент, синтезируемый в клетках Е. coli, и фермент, появляющийся в клетках Е. coli, инфицированных фагом Т4. Они различались по потребностям в кофакторах. ДНК-лигаза Е. coli в качестве кофактора требует дифосфопиридиннуклеотид, в то время как лигаза фага Т4 - аденозинтрифосфат. Кроме того, ДНК-лигаза фага Т4 в отличие от ДНК-лигазы Е. coli способна катализировать реакцию воссоединения двухцепочечных фрагментов ДНК с тупыми концами, т. е. фрагментов без перекрывающихся одноцепочечных комплементарных участков. Поэтому в настоящее время в генно-инженерных экспериментах предпочитают использовать ДНК-лигазу фага Т4, как более универсальный фермент.
ДНК-лигаза фага Т4 является мономерным полипептидом с молекулярной массой 68 кДа и катализирует образование фосфодиэфирной связи между прилегающими 5'-фосфатным (5'-р) и 3'-гидроксильным (3'-ОН) концами цепей ДНК. При этом возможны два типа реакций.
1. Лигирование липких концов:
Субстраты этой реакции - двухцепочечные молекулы ДНК с одноцепочечными, полностью комплементарными липкими концами. Частным случаем такой реакции является лигирование так называемого ника (nick) - разрыва в одной из нитей двухцепочечной ДНК.
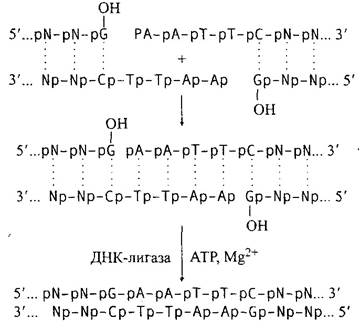
2. Лигирование тупых концов:
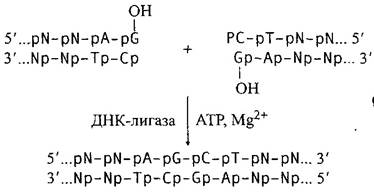
Таким образом, ДНК-лигаза фага Т4 обеспечивает ковалентное соединение любых двухцепочечных фрагментов ДНК, для которых имеется возможность состыковать 5'-р и 3'-ОН концы. Поэтому она является одним из важнейших ферментов, на использовании которых основаны современные методы рекомбинации молекул ДНК in vitro.








