 2015-07-21
2015-07-21 8253
8253ТЕМА 5. РАЗЛИЧНЫЕ ВИДЫ ПОДВИЖНОСТИ КОМПОНЕНТОВ МЕМБРАН
- Подвижность молекул липидов.
- Локализация и подвижность белков в бислое.
- Белок-липидные взаимодействия.
Подвижность молекул липидов.
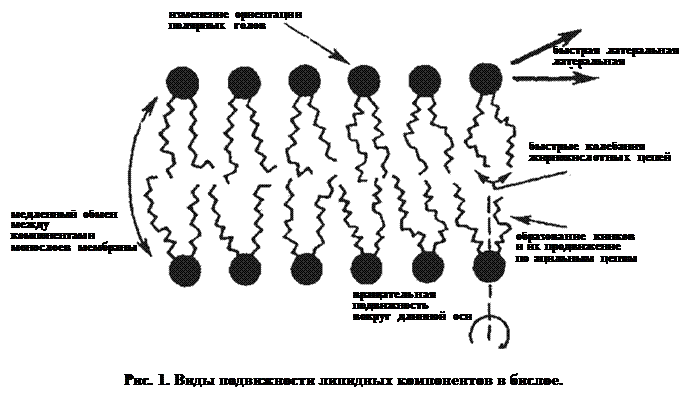
Молекулам в изотропной жидкости присущи разные виды подвижности: вибрационные колебания, вращение, трансляционные движения. В анизотропном бислое, напротив, молекулярная подвижность его компонент упорядочена. Различные типы подвижности проиллюстрированы на рис. 1.
Молекулы фосфолипидов способны к нескольким видам подвижности в бислое:
ü изменение ориентации полярных голов,
ü латеральное движение,
ü колебания ацильных цепей,
ü образование кинков и их перемещение вдоль ацильных цепей (в поперечном направлении),
ü ротационная подвижность (вращение вокруг длинной оси),
ü переход с одной стороны бислоя на другую (по типу флип-флоп),
ü выход из бислоя.
 |
Рассмотрим некоторые виды подвижности молекул фосфолипидов подробнее.
 1.1. Латеральное движение. Способность липидов перемещаться в мембране в латеральном (продольном) направлении показана многими экспериментами. Например, для суспензии яичного лецитина при 25ºС молекула преодолевает путь, равный 2,5 мкм за 1 сек. Таким образом, латеральная диффузия в упорядоченной мембране позволяет веществам перемещаться с относительно высокой скоростью. Она делает возможным образование липидных кластеров. Латеральная диффузия оказывается возможной даже при температуре кристаллического состояния. По-видимому, единственный механизм, который мог бы удовлетворительно объяснить этот факт, заключается в латеральной неоднородности мембраны, наличии дефектных зон (пустот), куда могут вытесняться молекулы из соседних упорядоченных областей.
1.1. Латеральное движение. Способность липидов перемещаться в мембране в латеральном (продольном) направлении показана многими экспериментами. Например, для суспензии яичного лецитина при 25ºС молекула преодолевает путь, равный 2,5 мкм за 1 сек. Таким образом, латеральная диффузия в упорядоченной мембране позволяет веществам перемещаться с относительно высокой скоростью. Она делает возможным образование липидных кластеров. Латеральная диффузия оказывается возможной даже при температуре кристаллического состояния. По-видимому, единственный механизм, который мог бы удовлетворительно объяснить этот факт, заключается в латеральной неоднородности мембраны, наличии дефектных зон (пустот), куда могут вытесняться молекулы из соседних упорядоченных областей.
1.2. Ротационная подвижность. Различные конфигурации молекул жирных кислот, возникающие при поворотах (вращении) вокруг единчной С-С связи, называют ротамерами, или конформерами, а изменение конформации молекулы за счет таких поворотов носит название транс-гош изомеризации. Вращательная подвижность молекул фосфолипидов, измеренная методом ЯМР, показала, что подвижность гидрофобных сегментов цепи повышается в направлении от сложноэфирной связи к метильной группе, т.е. к центру бислоя.
1.3. Трансмембранный переход «флип-флоп» типа. В липидных искусственных мембранах такие переходы осуществляются весьма медленно, например, полупериод перехода молекул холестерина с одной стороны бислоя на другую в липосомах из фосфатидилхолина занимает более 24 часов, однако, в принципе, такой переход возможен. Молекулы липидов не могут преодолеть липидный бислой в поперечном направлении путем перескока молекул с одной стороны бислоя на другую (флип-флоп), если в молекуле нет особых ферментов, известных под названием транслокаторов.
Липиды и в биологических мембранах с довольно большой частотой мигрируют с одной стороны мембраны на другую, то есть совершают «флип-флоп» переходы. Возможно, что гетерогенность липидного состава биологических мембран увеличивает вероятность «флип-флоп» перехода в природных мембранах. Одним из результатов этой гетерогенности является возможность образования гексагональной фазы (вывернутых везикул), кратковременное существование которых позволяет вовлекать молекулы липидов с одной стороны бислоя, а возвращать их на другую.
Следовательно, динамическое состояние бислоя с высокой подвижностью его компонентов определяется одновременно несколькими факторами. С одной стороны, это вращательная подвижность отдельных молекул фосфолипидов. Вблизи метильного конца она осуществляется для каждой молекулы независимо, но с приближением к полярной «голове» и возрастанием плотности упаковки (особенно начиная с 9 углеродного атома, ближе которого к поверхности бислоя не встречается цис -двойных связей) подвижность уменьшается.








