 2015-09-06
2015-09-06 4360
4360Очень широко распространён в организме человека. Мы регулируем обмен веществ при помощи изменения активности регуляторных ферментов. Этот механизм типичен для гормонов белково-пептидной природы, а также для гормонов, производных аминокислот.
Суть его сводится к следующему: в момент образования гормон-рецепторного комплекса внутри клетки начинает значительно увеличиваться количество молекул (вторичные мессенджеры, вторичные посредники) либо за счёт активации их синтеза, либо вследствии их поступления извне (Са2+). Это молекулы, которые могут запускать, как правило, каскад систем активации ферментов или изменять активность регуляторных ферментов. В конечном итоге мы наблюдаем изменение активности регуляторных ферментов, контролирующих строго определённую ветвь метаболизма.
Адреналин взаимодействует с рецептором, вследствие чего рецептор изгибается и часть рецептора, обращённая к цитоплазме, приближается G-белку (имеющего α, β, γ полипептидные цепи). G-белки могут быть позитивными (активируют ферменты) и негативными (тормозят ферменты). К α-цепи G-белка присоединяется молекула ГТФ (это возможно, когда существует гормон-рецепторный комплекс) и гидролизует его до ГДФ. Гидролиз только при приближении аденилатциклазы. При присоединении ГТФ к α-цепи происходит диссоциация трёх субъединиц G-белка (α, β, γ) и α-цепь G-белка присоединяется к ферменту аденилатциклазе, находящейся в неактивной форме. В ассоциированном виде этот фермент не работает. α-цепь выталкивает из аденилатциклазы

 ГДФ+Фн
ГДФ+Фн
ГТФ
Рисунок 2. Мембранно-внутриклеточный механизм (активация аденилатциклазы)
ингибирующую цепь, приводя к диссоциации самого фермента и активность аденилатциклазы увеличивается. ГТФ соединяется с α-цепью некоторый интервал времени, в тече ние которого аденилатциклаза активно работает. Затем идёт гидролиз ГТФ и G-белок вновь ассоциирует друг с другом и активность аденилатциклазы уменьшается. Но за это время внутри клетки образуется большое количество ц-3,5-АМФ. В этот момент времени наблюдаем усиление химического сигнала. 1 молекула гормона во временном интервале приводит к наработке сотни и тысячи молекул нуклеотидов (ц-3,5-АМФ, ц-3,5-ГМФ), следовательно, одновременно сотни и тысячи молекулы ферментов активируются, активируется протеинкиназная система, идёт модификация регуляторных ферментов (ковалентная модификация, например, гликоген).
Вторичные посредники существуют следующей разновидности:
- ц-3,5-АМФ и ц-ГМФ;
- кальциевая система;
- соединения белково-пептидной природы (белок IRS при действии на клетку инсулина).
Кроме того, в момент образования гормон-рецепторного комплекса внутриклеточно может быть активирована система кальция как вторичного посредника. Внутри клетки начинает расти концентрация Са2+. Причины увеличения его концентрации:
1. За счёт поступления Са2+ извне (рецептор-зависимая активация ионных каналов в момент образования гормон-рецепторного комплекса);
2. За счёт торможения выведения Са2+ из клетки;
3. За счёт выброса Са2+ из внутриклеточных депо (эндоплазматический ретикулум).
Очень часто в момент действия гормона на рецептор или образования гормон-рецепторного комплекса внутри клетки активируется фермент фосфолипаза С. Этот фермент начинает гидролизовать мембранные фосфолипиды, а конкретно инозитфосфатиды и в цитоплазму клетки начинают поступать ДАГи и инозитол-3-фосфат.
Поэтому часто считают, что система Са2+ как вторичного мессенджера включает 3 компонента:
1. относительно высокая концентрация Са2+;
2. ДАГи;
3. инозитол-3-фосфат.
Са2+ связывается с кальций-связывающим белком (кальмодулин), а комплекс кальций-кальмодулин обладает 2 действиями:
- непосредственно изменяет аткивность регуляторных ферментов;
- активирует каскад систем регуляции ключевых регуляторных ферментов.
ДАГи увеличивают сродство Са2+ к ферментам. Инозитол-3-фосфат повышает выброс Са2+ из внутриклеточных депо.
Цитозольный (прямой механизм) – типичен для стероидов, гормонов, которые легко проходят через клеточную мембрану или гормонов, гормон-рецепторные комплексы которых могут быть погружены через клеточные мембраны. Липофильные молекулы этой группы гормонов легко диффундируют сквозь плазматическую мембрану любых клеток, но только в клетках-мишенях они находят свой специфический рецептор, имеющий высокую степень сродства к гормону.
Гормон-рецепторные комплексы образуются в цитоплазме, рецепторы – внутри клетки. После образования гормон-рецепторного комплекса он движется в сторону ядра, затем довольно сложный механизм изменения активности генетического аппарата клетки. Как правило, активируется несколько генов, ответственных за синтез строго определённых регуляторных белков.
Известно, что активация образовавшегося гормон-рецепторного комплекса зависит от температуры и присутствия солей, при этом меняется величина, конформация и поверхностный заряд комплекса, и он приобретает способность связываться с хроматином. Гормон-рецепторный комплекс связывается со специфической областью ДНК и активирует либо инактивирует специфические гены. В результете избирательного воздействия на транскрипцию генов и синтез соответствующих мРНК происходит изменение содержания определённых белков, что сказывается на активности тех или иных процессов метаболизма. Далее начинается синтез белка и концентрация этого регуляторного фермента начинает плавно нарастать внутри клетки. Таким образом можно изменять активность метаболического пути (повышать синтез жирных кислот). Этот механизм реализуется медленно, через несколько часов, иногда – даже суток.
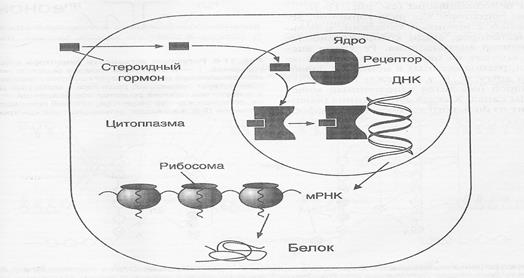
Рис. 3. Передача гормональных сигналов через внутриклеточные рецепторы (рецепторы стероидных гормонов могут находиться в цитоплазме и ядре).

