 2015-09-06
2015-09-06 16824
16824Мы можем рассматривать дрейф генов как один из факторов эволюции популяций. Благодаря дрейфу частоты аллелей могут случайно меняться в локальных популяциях, пока они не достигнут точки равновесия – утери одного аллеля и фиксации другого. В разных популяциях гены «дрейфуют» независимо. Поэтому результаты дрейфа оказываются разными в разных популяциях – в одних фиксируется один набор аллелей, в других – другой. Таким образом, дрейф генов ведет с одной стороны к уменьшению генетического разнообразия внутри популяций, а с другой стороны - к увеличению различий между популяциями, к их дивергенции по ряду признаков. Эта дивергенция в свою очередь может служить основой для видообразования.
В ходе эволюции популяций дрейф генов взаимодействует с другими факторами эволюции, прежде всего с естественным отбором. Соотношение вкладов этих двух факторов зависит как от интенсивности отбора, так и от численности популяций. При высокой интенсивности отбора и высокой численности популяций влияние случайных процессов на динамику частот генов в популяциях становится пренебрежимо малым. Наоборот, в малых популяциях при небольших различиях по приспособленности между генотипами дрейф генов приобретает решающее значение. В таких ситуациях менее адаптивный аллель может зафиксироваться в популяции, а более адаптивный может быть утрачен.
Как мы уже знаем, наиболее частым последствием дрейфа генов является обеднение генетического разнообразия внутри популяций за счет фиксации одних аллелей и утраты других. Мутационный процесс, напротив, приводит к обогащению генетического разнообразия внутри популяций. Аллель, утраченный в результате дрейфа, может возникать вновь и вновь за счет мутирования.
Поскольку дрейф генов – ненаправленный процесс, то одновременно с уменьшением разнообразия внутри популяций, он увеличивает различия между локальными популяциями. Этому противодействует миграция. Если в одной популяции зафиксирован аллель А, а в другой а, то миграция особей между этими популяциями приводит к тому, что внутри обеих популяций вновь возникает аллельное разнообразие.
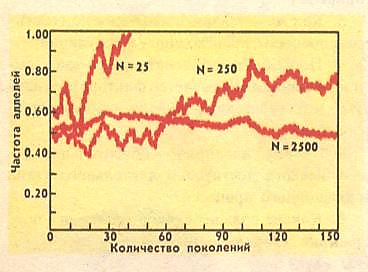
Рис. 3. N – число особей в популяции. Видно, что при 25 особях после 40-го поколения один аллель исчезает, при 250 – соотношение аллелей меняется, а при 2500 – остается близким к исходному.
Эффект бутылочного горлышка сыграл, по-видимому, очень значительную роль в эволюции популяций человека. Предки современных людей в течение десятков тысяч лет расселялись по всему миру. На этом пути, множество популяций полностью вымирало. Даже те, которые уцелели, часто оказывались на грани вымирания. Их численность падала до критического уровня. Во время прохождения через «бутылочное горлышко» численности частоты аллелей менялись по-разному в разных популяциях. Определенные аллели утрачивались полностью в одних популяциях и фиксировались в других. После восстановления численности популяций их измененная генетическая структура воспроизводилась из поколения в поколение. Эти процессы, по-видимому, и обусловили, то мозаичное распределение некоторых аллелей, которое мы сегодня наблюдаем в локальных популяциях человека. Ниже представлено распределение аллеля В по системе групп крови АВ0 у людей. Значительные отличия современных популяций друг от друга могут отражать последствия дрейфа генов, который происходил в доисторические времена в моменты прохождения предковых популяций через «бутылочное горлышко» численности.
Генетико-автоматические процессы, или дрейф генов, приводят к сглаживанию изменчивости внутри группы и появлению случайных, не связанных с отбором различий между изолятами. Именно это выявили наблюдения за особенностями фенотипов малочисленных групп населения в условиях, например, географической изоляции. Так, среди жителей Памира резус-отрицательные индивидуумы встречаются в 2—3 раза реже, чем в Европе. В большинстве кишлаков такие люди составляют 3—5% популяции. В некоторых изолированных селениях, однако, их насчитывается до 15%, т.е. примерно как в европейской популяции.
В крови человека имеются гаптоглобины, которые связывают свободный гемоглобин после разрушения эритроцитов, чем предотвращают его выведение из организма. Синтез гаптоглобина Нр1-1 контролируется геном Нр1. Частота этого гена у представителей двух соседних племен на Севере Южной Америки составляет 0,205 и 0,895, отличаясь более чем в 4 раза.
Примером действия дрейфа генов в человеческих популяциях служит эффект родоначальника. Он возникает, когда несколько семей порывают с родительской популяцией и создают новую на другой территории. Такая популяция обычно поддерживает высокий уровень брачной изоляции. Это способствует случайному закреплению в ее генофонде одних аллелей и утрате других. В результате частота очень редкогоаллеля может стать значительной.
Так, члены секты амишей в округе Ланкастер штата Пенсильвания, насчитывающей к середине девятнадцатого века примерно 8000 человек, почти все произошли от трех супружеских пар, иммигрировавших в Америку в 1770 г. В этом изоляте обнаружено 55 случаев особой формы карликовости с многопалостью, которая наследуется по аутосомно-рецессивному типу. Эта аномалия не зарегистирирована среди амишей штатов Огайо и Индиана. В мировой медицинской литературе описано едва ли 50 таких случаев. Очевидно, среди членов первых трех семей, основавших популяцию, находился носитель соответствующего рецессивного мутантного аллеля — «родоначальник» соответствующего фенотипа.
В XVIII в. из Германии в США иммигрировало 27 семей, основавших в штате Пенсильвания секту дункеров. За 200-летний период существования в условиях сильной брачной изоляции генофонд популяции дункеров изменился в сравнении с генофондом населения Рейнской области Германии, из которой они произошли. При этом степень различий во времени увеличивалась. У лиц в возрасте 55 лет и выше частоты аллелей системы групп крови MN ближе к цифрам, типичным для населения Рейнской области, чем у лиц в возрасте 28—55 лет. В возрастной группе 3—27 лет сдвиг достигает еще больших значений (табл. 1).
Рост среди дункеров лиц с группой крови М и снижение — с группой крови N нельзя объяснить действием отбора, так как направление изменений не совпадает с таковым в целом для населения штата Пенсильвания. В пользу дрейфа генов говорит также то, что в генофонде американских дункеров увеличилась концентрация аллелей, контролирующих развитие заведомо биологически нейтральных признаков, например оволосения средней фаланги пальцев, способности отставлять большой палец кисти (рис. 4).
Таблица 1. Прогрессивное изменение концентрации аллелей системы групп крови MN в популяции дункеров
| Возраст членов изолята, лет | Концентрация аллеля LM | LN |
| Более 55 От 28 до 55 От 3 до 27 | 0,55 0,66 0,735 | 0,45 0,34 0,265 |
На протяжении большей части истории человечества дрейф генов оказывал влияние на генофонды популяций людей. Так, многие особенности узкоместных типов в пределах арктической, байкальской, центрально-азиатской, уральской групп населения Сибири являются, по-видимому, результатом генетико-автоматических процессов в условиях изоляции малочисленных коллективов. Эти процессы, однако, не имели решающего значения в эволюции человека.
Рис. 4. Распространение нейтральных признаков в изолятедункеров штата Пенсильвания: а— рост волос на средней фаланге пальцев кисти, б— способность отставлять большой палец кисти
Последствия дрейфа генов, представляющие интерес для медицины, заключаются в неравномерном распределении по группам населения Земного шара некоторых наследственных заболеваний. Так, изоляцией и дрейфом генов объясняется, по-видимому, относительно высокая частота церебромакулярной дегенерации1 в Квебеке и Ньюфаундленде, детского цестиноза во Франции, алкаптонурии в Чехии, одного из типов порфирии среди европеоидного населения в Южной Америке, адреногенитального синдрома у эскимосов. Эти же факторы могли быть причиной низкой частоты фенилкетонурии у финнов и евреев-ашкенази.
Изменение генетического состава популяции вследствие генетико-автоматических процессов приводит к гомозиготизации индивидуумов. При этом чаще фенотипические последствия оказываются неблагоприятными. Гомозиготизация - это перевод гетерозигот в гомозиготы при близкородствснном скрещивании. Ч. Дарвин описывает явление, которое вполне можно объяснить дрейфом генов. «Кролики, одичавшие на острове Порто-Санто, близ о. Мадейры», заслуживают более полного описания*. Вместе с тем следует помнить, что возможно образование и благоприятных комбинаций аллелей. В качестве примера рассмотрим родословные Тутанхамона (рис.5) и Клеопатры VII (рис. 6), в которых близкородственные браки были правилом на протяжении многих поколений.
Тутанхамон умер в возрасте 18 лет. Анализ его изображения в детском возрасте и подписи к этому изображению позволяют предположить, что он страдал генетическим заболеванием — целиакией, которая проявляется в изменении слизистой оболочки кишечника, исключающем всасывание клейковины.
________________________________________________________
1церебромакулярная дегенерация, болезнь Тея – Сакса. Относится к группе наследственных липидных болезней мозга. На основании возраста начала болезни, клинических проявлений, картины глазного дна и данных биохимического исследования выделяют 5 форм амавротическойидиотии: врожденную, раннюю детскую, позднюю детскую, ювенильную и позднюю. Некоторые из этих форм отличаются и по характеру наследования.Характерный признак заболевания - диффузная дегенерация ганглиозных клеток во всех отделах нервной системы. Процесс распада ганглиозных клеток и превращения многих из них в зернистую массу - шафферовская дегенерация - является патогномоничным признаком амавротическойидиотии. Отмечаются также распад миелиновых волокон, особенно в зрительных и пирамидных путях, дегенеративные изменения глии. Врожденная форма - редкое заболевание. У ребенка уже при рождении отмечается микро- или гидроцефалия, параличи, судороги. Быстро наступает смерть. В мозговой ткани увеличено содержание ганглиозида Gm3.
Тутанхамон родился от брака Аменофиса III и Синтамоне, которая была дочерью Аменофиса III. Таким образом, мать фараона была его сводной сестрой. В могильном склепе Тутанхамона обнаружены мумии двух, по всей видимости мертворожденных, детей от брака с Анкесенамон, его племянницей.
Первая жена фараона была или его сестрой, или дочерью. Брат Тутанхамона Аменофис IV предположительно страдал болезнью Фрелиха и умер в 25—26 лет. Его дети от браков с Нефертити и Анкесенамон (его дочерью) были бесплодны. С другой стороны, известная своим умом и красотой Клеопатра VII была рождена в браке сына Птоломея Х и его родной сестры, которому предшествовали кровнородственные браки на протяжении по крайней мере шести поколений.
________________________________________________________________
* Это интересно
В 1418 или 1419 г. у ГонзалесаЗарко на корабле случайно оказалась беременная крольчиха, которая родила во время путешествия. Все детеныши были выпущены на остров. Кролики уменьшились почти на три дюйма в длину и почти вдвое в весе тела. По окраске кролик с Порто-Санто значительно отличается от обыкновенного. Они необычайно дики и проворны. По своим привычкам они более ночные животные. Производят от 4 до 6 детенышей в помете. Не удалось спарить с самками других пород". Примером воздействия дрейфа генов могут быть кошки о. Вознесения. Более 100 лет назад на острове появились крысы. Они расплодились в таком количестве, что английский комендант решил избавиться от них с помощью кошек. По его просьбе привезли кошек. Но они сбежали в отдаленные уголки острова и стали уничтожать не крыс, а домашнюю птицу и диких цесарок.
Другой комендант, чтобы избавиться от кошек, завез собак. Собаки не прижились — они ранили лапы об острые кромки шлака. Кошки со временем стали свирепыми и кровожадными. За столетие они отрастили себе почти собачьи клыки и стали сторожить дома островитян, ходить по пятам за хозяином и бросаться на посторонних.
Рис. 5. Родословная фараона XVIII династии Тутанхамона
Рис. 6. Родословная Клеопатры VII
Заключение и выводы:
Традиционно волны численности (жизни, популяционные) — присущие всем видам периодические и апериодические изменения численности особей в результате влияния абиотических и биотических факторов, воздействующих на популяцию, считаются "поставщиком" элементарного эволюционного материала.
Наилучшим доказательством значения дрейфа генов в микроэволюции
служит характер случайной локальной дифференциации в серии перманентноили периодически изолированных маленьких колоний. Дифференциация подобного типа многократно обнаруживалась в различных группах животных ирастений, популяции которых представляют собой систему колоний. Этадифференциация, если и не доказывает, то по крайней мере сильно склоняет кмнению о том, что дрейф генов играет важную роль в популяционных системах такого типа.
Использованная литература:
1. Гинтер Е.К Медицинская генетика: Учебник. – М.: Медицина, 2003. – 448 с.: ил
2. Грин Н., Стаут У., Тейлор Д «Биология» в 3 томах Москва «Мир» 2000г
3. Гуттман Б., Гриффитс Э., Сузуки Д., Кулис Т. Генетика. М.: ФАИР - ПРЕСС, 2004., 448 с
4. Жимулев И.Ф Генетика. Издательство Сибирского университета., 2007. – 480 с.:ил.
5. Курчанов, Н.А. Генетика человека с основами общей генетики. / Н.А. Курчанов. - СПб.: СпецЛит, 2006. - 174 с.
6. Мамонтов С.Г. Биология - М., 2004
7. Шевченко В.А., Топорнина Н.А., Стволинская Н.С. Генетика человека: Учеб.для студ. Высш. учеб. заведений. - М.: ВЛАДОС, 2002. - 240 с.9.
8. Ярыгин В.Н, В.И. Васильева, И.Н. Волков, В.В. Синелыцикова Биология. В 2 кн.: Учеб.для медиц. спец. Вузов М.: Высш. шк., 2003.— 432с.: ил.

