 2015-05-13
2015-05-13 2955
2955В результате формирования адаптаций достигается состояние адаптированности или соответствия строения, функций и поведения организмов занимаемым ими экологическим нишам. Экологическая ниша - это совокупность всех факторов среды, в пределах которых возможно существование вида в природе. Термин «экологическая ниша» предложен американским экологом Дж. Гринеллом (1917). Современная концепция экологической ниши сформировалась на основе модели экологической ниши американского эколога Дж. Хатчинсона (1957, 1965). Согласно этой модели, экологическую нишу можно представить как часть воображаемого многомерного пространства (гиперобъёма), отдельные измерения которого соответствуют факторам, необходимым для нормального существования вида. Расхождение экологических ниш разных видов путём дивергенции происходит чаще за счёт приуроченности к разным местообитаниям, потреблению различной пищи и разному времени использования одного и того же местообитания.
11.4. Специфика действия элементарных
эволюционных факторов в популяциях людей
11.4.1. Популяционная структура человечества
Термин «популяция» (или сообщество людей) используется в генетике и биологии человека для характеристики относительно стабильных и сравнительно изолированных групп людей. Популяция - это группа людей, занимающих определённую территорию и свободно вступающих в брак. Обычно под популяцией понимают как население города, района, так и определённой местности, используя этот термин в экологическом или географическом контекстах.
Для характеристики популяции важным является определение её как репродуктивного сообщества людей, обладающих общим генетическим фондом. Поэтому в формировании популяции людей главную роль играет не общность территории, а родственные связи между особями популяции. Демографическими показателями популяции людей служат размер, уровень рождаемости и смертности, возрастная структура, экономическое состояние, уклад жизни и т.п.
В популяции часто образуются более мелкие группировки людей, изолированных с точки зрения размножения (лишённых возможности свободно заключать брачные союзы в пределах популяции). Изолирующими факторами могут выступать географические (большое расстояние или препятствие к передвижению), но чаще всего факторы социального или религиозного порядка. При этом жители даже одного небольшого района могут часто образовывать ряд совершенно изолированных групп людей (изолятов) численностью до 1500 человек, но чаще всего численностью в несколько десятков особей. Закрепляющиеся гены имеют тенденцию ограничиваться именно этой группой (частота внутригрупповых браков в изолятах превышает 90%). Члены изолятов через 4 поколения (примерно через 100 лет) являются уже по крайней мере троюродными сибсами. Популяции людей численностью от 1500 до 4000 человек называют демами. Частота внутригрупповых браков в демах составляет 80-90%. В ранние периоды истории человечества расстояния и другие географические особенности служили наиболее устойчивыми барьерами на пути к смешению демов и изолятов, в чём кроется одна из причин широкой географической вариабельности человека как вида.
Строгой панмиксии в популяциях людей не существует: предпочтения в выборе брачного партнёра и социальные факторы препятствуют подлинно случайному заключению браков. Тем не менее существуют гены (например, определяющие группы крови), которые обычно редко принимаются во внимание при подборе брачных пар и распределяются среди жителей популяции так, как если бы подбор брачных пар происходил более или менее случайно.
Общая структура популяций человека - это структура, типичная для небольших долго существовавших изолятов, которая время от времени нарушалась в связи с миграциями, вторжением или смешением особей. Однако в настоящее время в связи с развитием транспорта круг возможных браков значительно расширился, замкнутость изолятов нарушается во всех частях Земли и, по-видимому, навсегда. Есть основания полагать, что широко распространённое в Европе увеличение роста, начавшееся в ХХ веке, частично связано с разрушением изолятов и уменьшением доли близкородственных браков (степени инбридинга).
11.4.2. Влияние мутационного процесса
на генетическую конституцию людей
В основе мутаций лежат наследуемые изменения генетического материала. В результате мутации возникает мутантная аллель гена или мутантная хромосома, обусловливающие появление мутантного признака. Мутации могут возникнуть в любой момент, но их появление более вероятно в делящейся, а не в покоящейся клетке (например, при гаметогенезе, во время мейоза). В генетическом отношении важны те мутации, которые возникают при гаметогенезе и наследуются особями потомства. Частота мутации для единичного локуса составляет в среднем 1:100000 половых клеток, однако у человека в целом, генотип которого насчитывает до 120000 (а возможно, и более) генов, мутация вовсе не редкое явление.
Мутагенами по отношению к человеку выступают не только естественные факторы (ультрафиолетовое излучение, температура, ионизирующее излучение, определённая химическая среда), но и факторы, производные научно-технического прогресса (рентгеновские излучения и другие физические факторы, синтетические смолы и другие химические вещества). На частоту мутаций у человека оказывает влияние возраст: вероятность рождения ребёнка, страдающего ахондропластической карликовостью, у пожилых супругов выше, чем у молодых. Некоторые гены Х-хромосомы мутируют в мужском организме чаще, чем в женском.
Генотип человека - это высокоинтегрированная система взаимодействующих генов (а также составляющих их элементов), и случайные изменения в её составе влияют на неё чаще всего отрицательно. Поэтому большинство мутантных генов оказываются вредными для человека.
В небольших популяциях людей мутантные гены могут сохраняться (фиксироваться) или утрачиваться случайным образом. В них хорошо выражен дрейф генов - изменение частоты генов в популяции в ряду поколений под действием чисто случайных (стохастических) факторов. На дрейф генов влияют такие факторы, как число индивидуумов, способных оставить потомство, и вариабельность в размере семьи. Структура популяции человека в прошлом создавала идеальные условия для дрейфа генов. Так, численность сообществ человека палеолита, очевидно, не превышала нескольких сот индивидуумов, именно такова численность сообществ современных охотников и собирателей - аборигенов Австралии.
В настоящее время общепризнанным является тот факт, что различия в частоте некоторых групп крови между близкими поселениями людей в отдалённых уголках земного шара возникли вследствие дрейфа генов: частоты генов в существующих в США небольших религиозных изолятах немецкого происхождения отличаются от соответствующих частот в исходной популяции в Германии и в соседних популяциях в США. Обобщённым примером влияния дрейфа генов на частоту аллелей является «эффект родоначальника». Он возникает, когда несколько семей выселяются на новую территорию и поддерживают высокий уровень брачной изоляции, порвав практически все связи с родительской популяцией. В этом случае в генофонде переселенцев из-за небольшой численности особей случайно закрепляются одни аллели и элиминируются другие. Так, среди кишлаков и других поселений Памира в одних популяциях резус-отрицательные индивидуумы составляют до 15%, а в других (таких большинство) - только 3-5%. Последний показатель для населения Памира в целом в 2-3 раза ниже, чем для населения Европы.
Последствиями дрейфа генов, очевидно, является неравномерное распределение некоторых наследственных заболеваний по группам населения земного шара. Так, высокая частота церебромакулярной дегенерации отмечена в Квебеке (Канада) и Ньюфаундленде; алкаптонурии - в Чехословакии; детского цестиноза - во Франции; адреногенитального синдрома - у эскимосов.
Существенное влияние на генофонды популяций людей оказывал фактор изоляции. Длительным проживанием в состоянии относительной географической и культурной изоляции объясняют, например, некоторые антропологические особенности представителей малых народностей: своеобразный рельеф ушной раковины бушменов, большую ширину нижнечелюстного диаметра коряков и ительменов, исключительное развитие бороды у айнов.
Сохранению высокого уровня генетической изоляции двух популяций (изолятов), существующих на одной территории, способствуют отличия по физическим признакам или образу жизни. Однако такие барьеры в последнее время исчезают, о чём свидетельствует, например, тот факт, что доля генов от белых людей возросла в настоящее время у американских негров до 25%, а у бразильских негров - до 40%.
11.4.3. Специфика действия естественного отбора
в человеческих популяциях. Популяционно-генетические эффекты действия систем отбора-контротбора
Одним из наиболее важных эволюционных факторов, изменяющих частоты аллельных генов в популяциях людей, является естественный отбор. Однако его давление в человеческих популяциях ослабело настолько, что отбор утратил значение как фактор видообразования. Это обусловлено возрастанием значения социальных факторов исторического развития человечества и постепенным ослаблением роли биологических факторов эволюции человека.
Однако за естественным отбором осталась функция стабилизации генофондов и поддержания наследственного разнообразия популяций людей. О действии на популяцию человека стабилизирующей формы естественного отбора свидетельствует, например, большая перинатальная смертность среди недоношенных и переношенных новорождённых. Направление отбора в этом случае определяется снижением общей жизнеспособности новорождённых.
Отрицательное действие отбора по одному локусу иллюстрирует наследование антигенов системы резус. 85% населения Европы имеет в эритроцитах антиген Rh и образует группу резус-положительных индивидуумов, остальные 15% населения составляют резус-отрицательные индивиды. Синтез антигена Rh контролируется доминантным аллелем D, который проявляется в гомозиготе (DD) и гетерозиготе (Dd). Поэтому резус-отрицательные люди являются рецессивными гомозиготами (dd). Если, например, мать резус-отрицательная (dd), отец резус-положительный (DD или Dd), то при беременности резус-положительным плодом (Dd) эритроциты плода могут проникнуть при нарушении плаценты в организм матери и иммунизировать его. При последующей (второй и т.д.) беременности резус-положительным плодом (Dd) антирезус-антитела, выработавшиеся в организме матери во время первой беременности, проникают через плаценту в организм плода и разрушают его эритроциты (эритробластоз). Развивается гемолитическая болезнь новорождённого, ведущим симптомом которой является тяжёлая анемия. В настоящее время успешно применяются различные способы борьбы с этой патологией (например, переливание новорождённому младенцу резус-отрицательной крови). В отсутствие медицинской помощи новорождённый с гемолитической болезнью часто погибал. Со смертью таких организмов (гетерозиготных по аллелю Dd) из популяции удаляется равное количество доминантных и рецессивных аллельных генов локуса «резус». Такой направленный против гетерозигот отбор приводит к уменьшению частоты более редкого (рецессивного, d) аллеля в европейской популяции. Теоретически в ряду из 600 поколений доля рецессивного гена может снизиться с 15% до 1%, на что потребуется около 15000 лет.
Несовместимость матери и плода характерна также для системы групп крови АВ0, которая, в отличие от резус-иммунизации, может сказаться уже на первом ребёнке. Однако эритробластоз в этом случае, как правило, выражен слабее, чем при Rh -несовместимости. Чаще всего АВ0 -несовместимость может приводить к увеличению числа спонтанных абортов. Отбор при этом идёт в основном против детей IAI0 и IB и I0 у матерей I0I0.
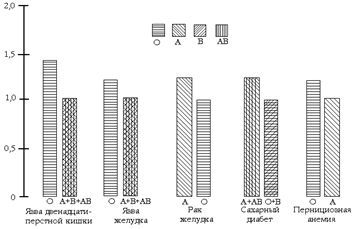
Для системы АВ0 существуют и другие селективные факторы: в настоящее время продемонстрирована ассоциация групп крови АВ0 с очень многими заболеваниями (рис. 153, А); например, лица с группой крови А чаще заболевают раком, тогда как лица, имеющие группу крови 0, более подвержены язве желудка и двенадцатиперстной кишки; а риск заболевания ревматизмом самый низкий среди лиц групп 0. Эти селективные факторы влияют на частоту аллелей, определяющих группы крови (рис. 153 Б, В).
Особой жёсткостью выделяется отбор, направленный против гомозигот: гомозиготы по многим рецессивным аутосомным заболеваниям обычно элиминируются, не достигнув репродуктивного возраста. Так, го-
|



мозиготы по аллелю серповидноклеточности эритроцитов (HbS/HbS) умирают от серповидноклеточной анемии в детском возрасте. Каждая такая смерть элиминирует из популяции аллели одного вида (рецессивные), что приводит к сравнительно быстрому снижению изменчивости по соответствующему локусу. Во многих популяциях людей частота аллелей аномальных гемоглобинов (включая аллель HbS) не превышает 1%. Отбор против гомозигот обусловлен также повышенной жизненной силой гетерозигот (явление гетерозиса).
Действию отбора, снижающего в генофондах некоторых популяций людей концентрацию определённых аллелей, может противостоять контротбор, который, наоборот, поддерживает частоту этих аллелей на достаточно высоком уровне. Проведённые в Северной Греции обследования показали, что больные серповидноклеточной анемией (гетерозиготы с генотипом HbS/HbA) болеют малярией в 13 раз реже, чем нормальные люди (HbA/HbA). Отрицательный отбор в отношении аллеля HbS перекрывается в этом случае мощным положительным отбором гетерозигот по аллелю HbS благодаря высокой жизнеспособности последних в очагах тропической малярии. Эта причина, действующая уже на протяжении нескольких столетий, обусловливает относительно низкую частоту гетерозигот по аллелю HbS среди североамериканских негров (8-9%) в сравнении с африканскими неграми (около 20%).
Генофонд популяций человека является результатом наложения многочисленных и разнонаправленных векторов отбора, обеспечивающего сохранение в каждом поколении сравнительно приспособленных к данным условиям генотипов. При этом с течением времени влияние отбора на генетическую структуру популяций людей снижается в основном благодаря успехам лечебной и профилактической медицины, а также социально-экономическим преобразованиям цивилизации.
11.4.4. Генетический полиморфизм и генетический
груз человечества. Генетический полиморфизм
Полиморфным признаком называют менделеевский (моногенный) признак, по которому в популяции присутствуют как минимум два фенотипа (и, следовательно, как минимум два аллеля), причём ни один из них не встречается с частотой менее 1% (т.е. не является редким). Эти два фенотипа (и, соответственно, генотипа) находятся в состоянии длительного равновесия. Наследственный полиморфизм создаётся мутациями и комбинативной изменчивостью. Часто в популяциях присутствует больше двух аллелей по данному локусу и, соответственно, более чем два фенотипа. Альтернативное полиморфизму явление - существование редких генетических вариантов, присутствующих в популяции с частотой менее 1%.
Первый полиморфный признак (система групп крови АВ0) был открыт в 1900 г. австрийским учёным К. Ландштейнером (1868-1943). В 1955 году с открытием методики электрофореза белков в крахмальном геле на примере гаптоглобина (сывороточного белка, связывающего гемоглобин) был выявлен самый простой вариант полиморфизма - полиморфизм белков.
К настоящему времени описано множество таких полиморфных признаков у человека:
1) сывороточные белки: церулоплазмин (2 аллеля - CPВ, СРС, а также более редкий аллель австралонегроидов - СРА); гаптоглобин (3 аллеля - НР1S, HP1F, HP2); иммуноглобины (4 аллеля и очень сложная система более редких аллелей);
2) поверхностные антигены эритроцитов (группы крови): АВ0 (4 аллеля: А1, А2, В, 0); секреция АВН (2 аллеля); антиген Келл (2 аллеля - K, k), антиген Льюис (2 аллеля - Lea, Leb); антиген резус (сложный комплекс аллелей);
3) ферменты эритроцитов: кислая фосфостаза-1 (3 аллеля); эстераза-D (2 аллеля); пептидаза-А (2 аллеля); аденозиндезаминаза (2+2 редких аллеля) и др.;
4) другие ферменты: сывороточная холинэстераза-1 (3 аллеля); алкогольдегидрогеназа (2 аллеля).
Различают наследственный и адаптационный полиморфизм. Наследственный полиморфизм создаётся мутациями и комбинативной изменчивостью. Адаптационный полиморфизм обусловлен тем, что естественный отбор благоприятствует разным генотипам из-за разнообразия условий среды в пределах ареала вида или сезонной смены условий. Например, в популяциях двухточечной божьей коровки (Adalia bipunctata) при уходе на зимовку преобладают чёрные жуки, а весной - красные особи. Это обусловлено тем, что чёрные жуки интенсивнее размножаются, а красные особи лучше переносят холод.
Разновидностью адаптационного полиморфизма является балансированный полиморфизм, возникающий в случаях, когда отбор благоприятствует гетерозиготным формам по сравнению с доминантными и рецессивными гомозиготами. В основе балансированного отбора может лежать сверхдоминирование - явление селективного преимущества гетерозигот (в том числе и над доминантными гомозиготами).
Различают следующие механизмы балансированного отбора: 1) обусловленность селективного преимущества гетерозигот их повышенной жизнеспособностью, основанной на явлении гетерозиса; повышение жизнеспособности происходит, очевидно, в результате взаимодействия аллельных генов во многих гетерозиготных локусах; 2) возникающие на основе гетерозиготности более редкие фенотипы могут получить в популяции селективные преимущества по двум причинам: а) самцы более редких (привлекательных) фенотипов имеют обычно повышенную конкурентоспособность в борьбе за самок и поэтому более значительный репродуктивный успех; б) хищники предпочитают более обычные для популяции фенотипические формы, не замечая редкие, возникшие на основе гетерозиготности;
3) любые мутации нарушают нормальную сбалансированность генотипа и фенотипа, поэтому они являются (чаще всего) вредными для организма и не могут быть сразу поддержаны отбором; в гетерозиготном же состоянии вредные мутации не проявляются, поэтому естественный отбор вначале благоприятствует не гомозиготным формам, несущим мутантный признак, а гетерозиготам, скрывающим этот признак от действия отбора.
Человечеству свойственен высокий уровень наследственного разнообразия. Кроме упомянутых выше многочисленных вариантов отдельных белков (простых признаков, прямо отражающих генетическую конституцию организма), люди отличаются друг от друга цветом кожи, глаз и волос, формой носа и ушной раковины, рисунком эпидермальных гребней на подушечках пальцев и другими сложными признаками. У людей не совпадают группы крови по системам эритроцитарных антигенов резус (Rh), АВ0 и другим. Известно более 130 вариантов гемоглобина, но лишь 4 обнаруживаются в нескольких популяциях в высокой концентрации: HbS (тропическая Африка, Средиземноморье), HbC (Западная Африка), HbD (Индия), HbE (тропическая и субтропическая зоны Юго-Восточной Азии).
Вариабельность распространения аллелей в популяциях людей зависит от действия элементарных эволюционных факторов особенно таких, как мутационный процесс и естественный отбор, а также дрейфа генов (генетико-автоматических процессов) и миграции особей. Межпопуляционным различиям в концентрации определённых аллелей способствует стабилизирующая форма естественного отбора. В основе стойкого сохранения в популяции людей одновременно нескольких аллелей одного гена лежит, как правило, отбор в пользу гетерозигот, который ведёт к состоянию балансированного полиморфизма.
Многие факторы отбора, действие которых создало современную картину распределения аллелей в популяциях людей, для большинства локусов не установлены, однако известна их экологическая природа. Экологическим проявлением отбора могут выступать инфекционные или паразитарные заболевания. В вышеприведённом описании заболеваемости серповидноклеточной анемией экологическим проявлением отбора гемоглобинов S, С и Е являлась малярия. Заболевания оказывают влияние также на распространение аллелей АВ0 (рис. 153). Так, люди с группой крови 0 более восприимчивы к чуме, чем люди с группой крови В. Однако вероятность заболеть раком желудка, ревматизмом, ишемической болезнью сердца, холециститом, желчнокаменной болезнью для них примерно на 20% ниже, чем для лиц с группой крови А.
Полиморфизм человечества по отдельным локусам мог быть унаследован от далёких предков. Так, полиморфизм по таким системам групп крови, как АВ0 и резус, обнаружен у человекообразных обезьян. Учитывая плохие экономические и гигиенические условия жизни основной массы населения Земли на протяжении значительной части истории человечества, можно полагать, что наследственный полиморфизм благоприятствовал выживанию в разных экологических ситуациях и способствовал расселению людей. Вклад в наблюдаемое сейчас распределение аллелей внесли массовые миграции населения и сопутствующая им метисация: смешение больших контингентов людей разной расовой принадлежности имело место в Восточной Африке, Индии, Индокитае, Южной и Центральной Америке.
Генетический полиморфизм служит основной межпопуляционной и внутрипопуляционной изменчивости людей. Эта изменчивость, в частности, проявляется: 1) в разной степени предрасположенности людей к определённым болезням; 2) неравномерном распределении по планете некоторых заболеваний; 3) неодинаковой тяжести их течения в разных человеческих популяциях; 4) индивидуальных особенностях течения патологических процессов; 5) различиях индивидуальной реакции на одно и то же лечебное воздействие. Генетический полиморфизм создаёт серьёзные трудности в решении проблемы пересадок тканей и органов.
Генетический груз. В пределах ареала какого-либо вида условия среды различны. Это становится причиной различного направления отбора в разных частях ареала. Поэтому естественный отбор действует на популяцию (вид) одновременно по многим направлениям, а его конечный результат зависит от соотношения интенсивности разных векторов отбора и контротбора. Результатом многовекторного действия отбора в пределах ареала является поддержание в разнообразном состоянии и одновременная относительная стабилизация генофонда популяции.
Генетически разнородная популяция более жизнеспособна, всегда получая селективное преимущество при часто наблюдаемых колебаниях условий (параметров) среды. В генофонде такой популяции накапливается (особенно в гетерозиготном состоянии) большой объём резервной наследственной изменчивости. Такая популяция в эволюционном плане более пластична, может более гибко реагировать на изменения условий среды, приспосабливаясь к ним. Однако в такой популяции неизбежны особи, менее приспособленные к данным условиям среды. В каждый отдельно взятый момент жизнеспособность такой популяции ниже уровня, который был бы достигнут при наличии в популяции только наиболее подходящих для данных условий генотипов. Часть наследственной изменчивости популяции, которая определяет появление менее приспособленных к данным условиям особей, называется генетическим грузом. Иными словами, генетический груз - это величина, на которую приспособленность реальной популяции отличается от приспособленности идеальной для данных условий популяции, состоящей из «лучших» (при данном генофонде) генотипов. Источником генетического груза служат мутационные и сегрегационные процессы.
Вместе с тем, поддерживая генетическое разнообразие и, следовательно, эволюционную пластичность популяций, генетический груз представляет собой одновременно генотипический резерв эволюции. При изменении направления естественного отбора особи с уклоняющимися от доминирующего фенотипами (генотипами) обеспечивают выживание и эволюционную перспективу целостной популяции. Следовательно, генетический груз - это своеобразная плата за экологическую пластичность и эволюционную перспективу. Генетический груз - неизбежное следствие генетического полиморфизма.
Различают следующие виды генетического груза:
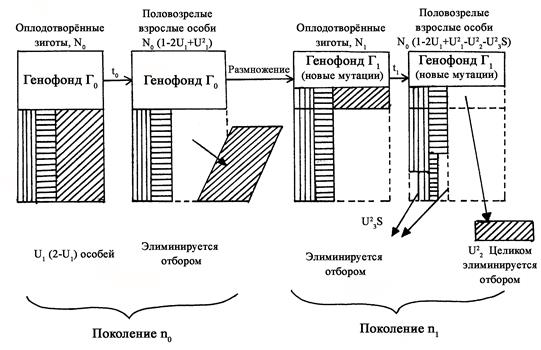
а) мутационный груз - обусловлен возникновением в популяции мутантных аллелей (рис. 154): поскольку отбор направлен против этих аллелей, их частота в популяции невелика и она поддерживается благодаря повторному возникновению (мутационному давлению);
б) сегрегационный груз - возникает в результате выщепления гетерозиготными родителями менее приспособленных гомозиготных потомков; в связи с тем, что значительная часть мутантных аллелей оказывает в гетерозиготном состоянии положительное действие (эффект сверхдоминирования), то гетерозиготы (а следовательно, и вредные мутации) могут поддерживаться в ряду поколений;
В) субституционный груз - возникает при изменении адаптивной ценности особей и сохраняется в популяции до тех пор, пока другой аллель не заместит потерявший адаптивную ценность первый аллель.
Летальный эквивалент. Генетический груз снижает в целом реальную приспособленность популяций людей к данным условиям. Для оценки бремени генетического груза человечества используют понятие летальных эквивалентов. Их число у отдельных людей составляет от 3-х до 8-ми. Это значит, что преобладающие количество неблагоприятных аллелей, которое имеется в генотипе каждого человека, оказывает суммарное вредное действие, эквивалентное действию 3-8 рецессивных аллелей, каждый из которых вызывает в гомозиготном состоянии смерть индивидуума до наступления репродуктивного возраста.
Из-за наличия неблагоприятных аллелей и их сочетаний примерно половина зигот, образующихся в каждом поколении людей, в биологическом
|
плане несостоятельна, т.к. не участвует в передаче генов следующему поколению: около 15% организмов гибнет до рождения, 3% - при рождении, 2% - сразу после рождения, 3% людей умирает, не достигнув половой зрелости, 20% лиц не вступает в брак и 10% образующихся браков бездетны.

