2015-10-13
2015-10-13 734
734Происхождение эвкариот
При рассмотрении последующих этапов эволюции жизни необходимо ответить на несколько основных вопросов.
1.Чем было обусловлено дальнейшее усложнение организации живых организмов, которое является одним из определяющих признаков прогрессивной эволюции.
2.Когда происходили изменения, которые привели к появлению более сложных живых организмов
3.Какие механизмы обеспечили более сложную организацию организмов в ходе эволюции и чем подтверждается их существование.
Из предыдущего изложения ясно, что биосфера древней Земли была создана прокариотами. Именно они были активными участниками сформировавшихся на Земле глобальных циклов веществ и энергии и их стабилизаторами. Наиболее интегрированными системами сообществ прокариот, были бактериальные маты, создавшие повсеместно строматолиты, которые практически неизменными сохранились до наших дней. Однако на определённом этапе истории Земли в составе биосферы появились организмы, вначале одноклеточные, которые существенно отличались от прокариот более сложным строением клетки – эвкариоты. Наиболее существенные различия прокариот и эвкариот отражены в таблице 3.1 и на рисунке 3.1..
Таблица 3.1. Основные различия прокариот и эвкариот
| Прокариоты | Эвкариоты | |
| Группы организмов | Бактерии, архебактерии, цианобактерии | Простейшие, грибы, растения, животные |
| Размеры клетки | Мелкие, обычно 1 – 10 мкм (до 60 мкм) | Крупные, обычно 30 – 100 мкм |
| Метаболизм и фотосинтез | Анаэробные или аэробные | Аэробные, редкие случаи анаэробиоза (дрожжи) вторичны |
| Подвижность | При помощи жгутиков из белка флагеллина | Реснички и жгутики – ундулиподии - из белка тубулина |
| Клеточная мембрана | Единственная молекула мукопептида из аминокислот, углеводов и липидов | Двухслойная мембрана из слоёв липидов, пронизанная молекулами белков |
| Органеллы | Мембранные органеллы отсутствуют, рибосомы мелкие | Митохондрии, хлоропласты, аппарат Гольджи и др., рибосомы крупные |
| Генетический аппарат | Петля ДНК в цитоплазме | ДНК в хромосомах, окружённых ядерной мембраной |
| Размножение клеток | Деление надвое | Митоз или мейоз с образованием веретена |
| Клеточное строение | Клетки одного типа | Сложные клеточные циклы или многоклеточные и многотканевые |
| Движение цитоплазмы | Отсутствует | Сложные движения, упорядоченные с помощью сократимых элементов цитоскелета |
Важно отметить, что эти две группы чётко отграничены друг от друга, никаких промежуточных форм не существует. И есть определённые основания полагать, что их никогда не существовало в истории Земли.
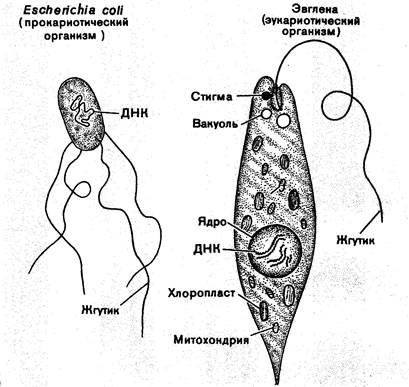
Рисунок 3.1. Основные различия клетки прокариот и эвкариот
Появление эвкариот – событие в истории Земли не менее значимое, чем возникновение жизни. Прокариоты оказались неспособными к дальнейшему усложнению своей организации. Особенности реализации генетических программ и изменений в них в результате мутаций обеспечивали крайне расточительный способ адекватного реагирования на изменение условий существования – все непригодные особи сразу отбраковывались. Это компенсировалось высокой скоростью размножения и способностью горизонтальной передачи полезных свойств даже между различными группами прокариот. Но путь прогрессивной эволюции для них был закрыт. Настоящая многоклеточность у них так и не возникла. Причиной видимо служили особенности строения мембраны клеток и отсутствие механизма митоза, позволяющего обеспечить точную ориентацию клеток в пространстве.
Основные особенности эвкариот, определившие их эволюционную перспективность, включают следующие признаки.
· Обособление клеточного ядра от цитоплазмы – обеспечивает оптимальные условия для хранения и защиты информации и функционирования клетки.
· Обособление ядра позволяет осуществлять более совершенную регуляцию генома, благодаря чему возникли сложные механизмы приспособительной (модификационной) изменчивости, а, следовательно - достигнут более высокий уровень приспособляемости.
· Возможность формировать различные типы клеток при неизменном геноме позволила одноклеточным эвкариотам в дальнейшем сформировать:
- сложные клеточные циклы;
- половое размножение;
- многоклеточность.
· Обособление ядра предоставило возможность развить более совершенные системы защиты и репарации (восстановления) ДНК. Это резко снизило частоту мутирования, что обеспечило эвкариотам преимущество в условиях жёсткого ультрафиолетового излучения, которое было характерно в условиях бескислородной атмосферы архея и протерозоя в отсутствие озонового экрана. Такой механизм защиты мог значительно увеличить зону обитания первых эвкариот, позволив им освоить верхние лучше освещённые слои воды.
В 1999 г. В отложениях возраста 2,7 млрд. лет были обнаружены остатки стероидных соединений, характерных только для мембран эвкариотических клеток. Этот период – переход от верхнего архея к раннему протерозою – характеризовался крайне нестабильной геологической обстановкой, что было связано с формированием железного ядра Земли, резкой активизацией тектонических процессов, образованием единого суперконтинента (Мононгеи). В этот период происходили и другие перемены – за счёт изменений газового состава атмосферы (падение концентрации углекислоты и метана – основных парниковых газов) снизилась температура, что вызвало первые оледенения. Резко повысилась pH морской воды – от 3–5 в конце архея до 7–8 в начале протерозоя. От этих изменений страдали теплолюбивые прокариотные сообщества, в период с 2,5 по 2,3 млрд. лет заметно снизилось количество строматолитов. Колебания условий среды оказались слишком большим испытанием для адаптивной стратегии прокариот (быстрое мутирование, обмен генами и отбор устойчивых клонов) и они оказались менее конкурентоспособными по сравнению с эвкариотами. Последние могли возникнуть и раньше, но именно в этих условиях они становятся заметным компонентом биосферы и обнаруживаются в ископаемом состоянии.
Существует несколько гипотез происхождения эвкариот. Наиболее аргументированной и подтверждаемой рядом убедительных доказательств является гипотеза симбиотического происхождения эвкариот, согласно которой они возникли в результате симбиотического взаимодействия и последующего объединения различных прокариот в единый одноклеточный организм. Основание для такого предположения впервые было представлено в работе физиолога А.С.Фаминцына о строении лишайников, которые оказались сообществом гриба и водоросли. Позднее Фаминцын прямо сформулировал предположение о симбиотической природе растительных и животных клеток. Эти взгляды развивались и другими отечественными биологами – К.С.Мережковским, Б.М.Козо-Полянским. Однако только в 60-е годы прошлого века эта гипотеза получила новый импульс благодаря работам Л.Маргелис.
Под симбиозом понимают ассоциации между особями различных видов, сохраняющиеся на протяжении значительной части жизненного цикла. Различают мутуализм – симбиоз взаимно полезный для партнёров, и комменсализм – симбиоз безвредный для них.
Аргументы в пользу симбиотического происхождения эвкариот можно разделить на несколько основных групп.
К первой группе относятся свидетельства широкого распространения явлений симбиоза среди живых организмов. В известном смысле симбиоз является обязательным условием существования большинства организмов самых различных уровней организации. Степень симбиотического слияния может варьировать так сильно, что в ряде случаев трудно провести границу между, например, внутриклеточными симбионтами и органеллами эвкариотической клетки.
Симбиоз возникает в случаях, когда совместное существование оказывается в определённых условиях более выгодным, чем раздельное. Это состояние может быть обратимым, если условия среды делают более эффективной самостоятельную жизнь. Так, лишайники могут при определённых изменениях условий (влажность, температура) распадаться на гриб и водоросль. Однако в ряде случаев только симбиотическая ассоциация позволяет реализовать функции, которые у каждого члена сообщества по отдельности невозможны. Например, симбиотические ассоциации бактерий на корнях бобовых (клубеньки) фиксируют атмосферный азот, хотя каждый из партнёров в отдельности такой способности лишён.
Обязательным условием стабильности симбиоза являются одинаковые скорости роста партнёров, что требует наличия систем координации этого процесса. Такая координация, также как согласование более частных функций, предполагает существование определённых форм контроля, в том числе и генетических механизмов. Чаще всего это происходит путём утраты ряда генов внутриклеточными симбионтами и контроля соответствующих функций генами ядра клетки-хозяина или передачи генов в ядро клетки - хозяина. При длительном сосуществовании внутриклеточные симбионты утрачивают некоторые элементы своей структуры, оказывающиеся ненужными внутри клетки-хозяина. У водорослей-эндосимбионтов часто происходит редукция клеточной стенки и они становятся похожими на пластиды. В ряде случаев происходит редукция некоторых систем организма хозяина. Например, погонофоры, обитающие в зонах действия «чёрных курильщиков» - местах выхода горячих гидротермальных источников на дне океана, вообще лишены пищеварительной системы. Питание этих своеобразных животных обеспечивается бактериями - хемоавтотрофами, которые находятся в специальных камерах внутри животного и снабжают своего хозяина органикой, вырабатываемой в процессе хемосинтеза.
Очень широко распространены симбиозы между светящимися бактериями и различными морскими животными. Степень связи между симбионтами значительно варьирует - от автономии при случайном попадании бактерий в кишечник хозяина до внутриклеточного симбиоза у одного из оболочников. Такие ассоциации всегда дают организму-хозяину явное преимущество. В ряде случаев, как, например, у рыб, развиваются даже специальные органы, в которых находятся бактерии. Эти органы могут быть снабжены специальными ширмами, позволяющими регулировать длительность световых импульсов для привлечения полового партнёра при размножении, освещении добычи или отпугивания хищников. В ряде случаев эти бактерии не удаётся культивировать вне организма хозяина, что свидетельствует об утрате ими ряда метаболических путей.
Для понимания процессов перехода от симбиотической связи к органеллам эвкариот наиболее интересны примеры внутриклеточного симбиоза с участием прокариот, поскольку только они могли быть участниками симбиоза, который привёл к возникновению эвкариотической клетки. Для таких случаев установлено, что в процессе совместного существования внутриклеточный симбионт может утратить значительную часть своих генов. Примером служит жгутиконосец Cyanophora paradoxa, клетки которого содержат цианеллы – редуцированные цианобактерии, содержащие не более 10% ДНК, которым располагают свободноживущие цианобактерии. Участниками симбиоза могут быть даже отдельные органеллы клеток. Так, оказалось, что в клетках некоторых моллюсков могут длительное, до нескольких месяцев, время культивироваться пластиды водорослей, используемых моллюсками в пищу. Интересен случай множественного симбиоза, когда он состоит из пяти различных организмов. В кишечнике австралийского термита обитает жгутиконосец, обеспечивающий переработку растительной клетчатки. В свою очередь, он является обладателем ещё трех бактерий – симбионтов, которые находятся на его поверхности, и одного внутриклеточного симбионта, который выполняет функции митохондрий, отсутствующих у этой инфузории. Спирохеты – подвижные бактерии, покрывающие поверхность инфузории, совершают синхронные движения и обеспечивают плавное поступательное движение инфузории вдоль стенки кишечника. Есть основания полагать, что согласованность их движений координируется управляющими сигналами клетки – хозяина.
Инфузория Paramecium bursaria, представляющая симбиотический комплекс инфузории и водоросли – хлореллы в природных условиях, неспособна реализовать половой цикл, если её лишить хлорелл – симбионтов, что свидетельствует о необратимых генетических изменениях у партнёров.
Все эти данные подсказывают возможные пути перехода симбиотических взаимодействий различных прокариот в связи, которые наблюдаются между эвкариотической клеткой и её органеллами. Но для подтверждения такого пути нужны факты, отражающие различия между эвкариотической клеткой – хозяином с одной стороны, и сходство органелл клетки с прокариотами – с другой.
Наиболее убедительные доказательства симбиотического происхождения органелл эвкариотических клеток получены в отношении митохондрий и пластид. Рассмотрим их по порядку.
1.Митохондрии сохранили целый ряд признаков, характерных для свободноживущих прокариот:
- кольцевую ДНК, которая синтезируется на всём протяжении жизненного цикла и поровну распределяется между дочерними митохондриями при их делении пополам;
- состав ДНК митохондрий более сходен с ДНК прокариот, чем с ядерной ДНК эвкариот;
- появление митохондрий сделало возможным синтез производных стероидов и ряда других вторичных метаболитов;
- митохондрии независимо распределяются между дочерними клетками с помощью различных механизмов;
- рибосомные и транспортные РНК митохондрий отличаются от ядерных;
- рибосомы митохондрий отличаются по структуре и составу белков и ферментов от рибосом цитоплазмы клетки.
Наиболее вероятным предком митохондрий могла быть аэробная бактерия - гетеротроф, содержащая системы для полного окисления углеводов до СО2 и Н2О.
2.К признакам, свидетельствующим о возможном прокариотическом происхождении пластид растительных клеток, можно отнести следующие:
- пластиды всегда образуются путём деления исходных пластид, у некоторых водорослей и мхов пластиды делятся регулярно при каждом клеточном делении;
- пластиды содержат ДНК, информационную РНК и системы белкового синтеза;
- при удалении пластид с помощью различных методов клетки навсегда утрачивали способность к их образованию;
- ДНК пластид имеет кольцевую форму подобно ДНК прокариот;
- РНК рибосом из пластид неродственна РНК рибосом из цитоплазмы той же клетки, но обладает значительно большим сходством с РНК рибосом прокариот;
- передача пластид зиготе только через женскую клетку определяется наличием у неё цитоплазмы, содержащей пластиды.
Наиболее вероятными предшественниками пластид были фотосинтезирующие прокариоты, выделяющие в окружающую среду свободный кислород и сходные с современными цианобактериями и зелёными прокариотами Prochloron - пигменты последних имеют тот же состав, что и у зелёных растений.
В настоящее время предполагают, что митохондрии вошли в состав органелл очень рано, даже раньше образования клеточного ядра. Это подтверждается сходством молекулярной структуры митохондрий у всех основных групп эвкариот. Пластиды возникли в ходе эволюции позднее.
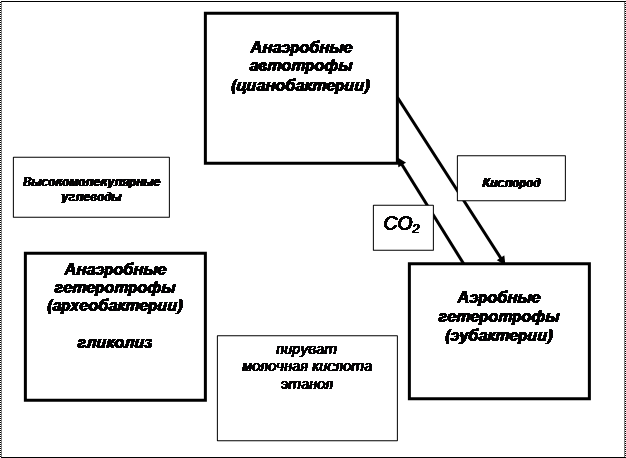
Условия, в которых происходило становление эвкариот, связывают с существованием устойчивых прокариотических сообществ, которыми были, скорее всего, цианобактериальные маты. Симбиотические взаимоотношения составляющих их прокариот создавали ассоциации, в которых были хорошо отрегулированы взаимовыгодные процессы обмена продуктами жизнедеятельности членов сообщества. Предполагают, что в его состав входило минимум три компонента (Рис.3.2)


 некоторые виды
некоторые виды
 | |||||
 | |||||
 | |||||
Рис. 3.2. Минимальный состав сообщества прокариот – родоначальников
эвкариотической клетки
· Анаэробные автотрофы (цианобактерии), которые производили высокомолекулярные углеводы и выделяли в окружающую среду свободный кислород.
· Анаэробные гетеротрофы (вероятно – археобактерии), путем гликолиза сбраживавшие углеводы с образованием промежуточных органических соединений – пирувата, молочной кислоты, спирта.
· Аэробные гетеротрофы (эубактерии), производившие кислородное окисление продуктов, выделяемых археобактериями, с образованием конечных низкоэнергетических продуктов – углекислоты и воды.
Между членами этого симбиотического сообщества происходил взаимовыгодный обмен продуктами жизнедеятельности, что со временем могло делать эти отношения всё более тесными. Цианобактерии и археобактерии избавлялись от излишков токсичного для них кислорода, а также от конечных продуктов бескислородного энергетического обмена; археобактерии и аэробные эубактерии получали необходимую им для питания органику, а последние – еще и кислород как мощный окислитель, позволяющий с высокой эффективностью утилизировать органические соединения. Структура этого симбиоза – не что иное как усовершенствованный рассмотренный ранее цианобактериальный мат, совместная эволюция элементов которого превратила его в своего рода квазиорганизм. Поэтому можно предположить, что многие из механизмов эвкариотической клетки начали вырабатываться задолго до объединения её компонентов под одной клеточной мембраной. Способность горизонтального обмена генетической информацией, свойственная прокариотам, при их объединении под общей оболочкой и возникновении неизбежного дублирования ряда функций могла облегчить передачу многих генов от симбионтов ядру клетки и редукцию генов органелл.
В связи с формированием эвкариотической клетки путём слияния клеток прокариот возникают два вопроса: какой тип прокариот был клеткой-хозяином и какие механизмы обеспечили проникновение через мембрану клетки-хозяина. Известно, что по крайней мере современные прокариоты не способны к фагоцитозу.
Наибольшие шансы на роль клетки-хозяина имели, по современным представлениям, архебактерии. В их строении имеется ряд особенностей, сходных с эвкариотами:
· экзон-интронная структура генома;
· сходные механизмы репликации, репарации, транскрипции и трансляции;
· наличие механизмов защиты генетической информации от внешних воздействий.
Сохранение этих свойств у эвкариотического организма безусловно обеспечило высокую степень защищённости от неблагоприятных условий для всех органелл клетки.
Проникновение через мембрану клетки-хозяина было возможным для прокариот, представители которых существуют и в настоящее время – это некоторые хищные и паразитические бактерии. Последующий переход от паразитизма к симбиозу также известен и описан в литературе.
Дальнейшая эволюция эвкариот связана с появлением полового процесса (коньюгации), который вероятно появился как способ противодействия неблагоприятным условиям. У современных простейших половой процесс возникает именно в такие периоды, при преобладании бесполого размножения в благоприятные сезоны. Половой процесс, способствуя рекомбинации генетического материала, увеличивает изменчивость, что способствует выживанию при резком изменении условий обитания за счёт увеличения разнообразия.
Последствием полового процесса неизбежно стало появление диплоидной стадии и мейоза, а затем рецессивной изменчивости. Эти процессы сопровождались возникновением хромосом и механизма обмена генами между ними – кроссинговера.
Результатом всех этих новообразований стало резкое повышение приспособляемости эвкариот и их сообществ. Однако, оставаясь на уровне одноклеточных организмов, они могли эволюционировать лишь по пути усложнения организации клетки и создания сложных клеточных циклов. Наиболее радикальным путём дальнейшего повышения уровня организации стало возникновение многоклеточности, а затем – многотканевых организмов, что привело к огромному разнообразию жизненных форм и всё возрастающей эффективности функционирования приспособительных механизмов.