 2015-10-13
2015-10-13 1866
1866представлена
· клеточным
· гуморальным иммунитетом.
Различают в зависимости от механизма уничтожения антигена
При клеточном иммунитете эффекторными клетками являются цитотоксические Т-лимфоциты, или лимфоциты киллеры (убийцы), которые непосредственно участвуют в уничтожении чужеродных клеток других органов или патологических собственных (например, опухолевых) клеток. Среди лимфоцитов в крови, кроме вышеописанных Т-ц, выполняющих функцию киллеров, имеются так называемые натуральные киллеры (Нк), которые также участвуют в клеточном иммунитете. Они образуют первую линию защиты против чужеродных клеток, действуют немедленно, быстро разрушая клетки. Нк в собственном организме разрушают опухолевые клетки и клетки, инфицированные вирусом. Тц образуют вторую линию защиты, так как для их развития из неактивных Т-лимфоцитов требуется время, поэтому они вступают в действие позже.
Клеточный иммунный ответ формируется при трансплантации органов и тканей, инфицировании вирусами, злокачественном опухолевом росте. В клеточном иммунитете участвует Тц (Тк), реагирующий с антигеном в комплексе с гликопротеинами МНС 1 класса в плазматической мембране клетки-мишени. Цитотоксическая Т-клетка убивает клетку, инфицированную вирусом, в том случае, если она узнает с помощью своих рецепторов фрагменты вирусных белков, связанные с молекулами МНС класса I на поверхности зараженной клетки.
Связывание Тц с мишенями ведет к высвобождению цитотоксическими клетками порообразующих белков, называемых перфоринами, которые полимеризуются в плазматической мембране клетки-мишени, превращаясь в трансмембранные каналы. Как полагают, эти каналы делают мембрану проницаемой, что способствует гибели клетки.
При гуморальном иммунитете имеются эффекторные клетки, которые синтезируют и выделяют в кровь антитела. Антитела — это сложные белки, синтезируемые В-лимфоцитами и плазмоцитами, способные специфически соединяться с соответствующими антигенами (например, с бактериальными) и обезвреживать их. Обнаружение антител в глобулиновой фракции белков крови обу 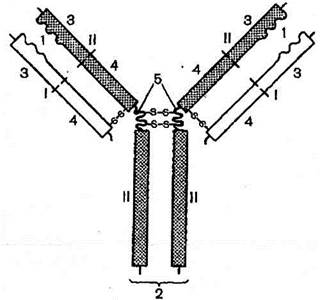 словило их название — иммуноглобулины
словило их название — иммуноглобулины
Рис. 223. Строение молекулы антитела (схема по Б.Альбертсу и др.).
I — легкие цепи (L);
II - тяжелые цепи (Н);
1 — антигенсвязывающие участки (РаЬ-фрагменты) Н- и L-цепей;
2 — кристаллизующийся фрагмент (Fc-фрагмент) Н-цепей;
3 — вариабельные области (V) Н- и L-цепей;
4 — константные области (С) Н- и L-цепей;
5 — дисульфидные мостики.
Молекула антитела имеет форму V и состоит из четырех полипептидных цепей — двух идентичных тяжелых цепей — Н- цепей и параллельно расположенных двух идентичных легких цепей — L-цепей, соединенных дисульфидными мостиками. Каждая Н- и L-цепь иммуноглобулиновой молекулы имеет вариабельные области V, располагающиеся на обоих ветвях Н- и L-цепей, и постоянные / константные области (С). В вариабельных областях находятся антигенсвязывающие участки — два РаЬ-фрагмента - места распознавания и связывания антигена. Постоянные области находятся в Fc-фрагменте, образованном лишь Н-цепями. Эти области обеспечивают связывание компонентов комплемента и/или клеточных рецепторов.
Выявлено 5 типов тяжелых цепей (|^,у, а, А, Е) и 2 типа легких цепей (к, X), различные сочетания которых обеспечивают образование множества разновидностей антител с уникальными участками связывания антигенов.
Антитела в высоких концентрациях находятся в крови и лимфе, а также в жидких секретах (молоко, слезы, пот, вагинальный секрет, секрет предстательной железы и др.).
Выявлено несколько классов иммуноглобулинов —
· Ig G,
· Ig M,
· Ig A,
· Ig D
· Ig E,
Ig G - Циркулирующее антитело в крови и лимфе. Соединяется с антигенами и активизирует систему комплемента (опсонизация) и лизис клеток. Их содержание повышается при инфекциях и снижается при недостатке гуморального иммунитета.
Ig M - Менее специфичен к антигенным детерминантам, но активен в реакции комплемента, опсонизации и нейтрализации токсических агентов. Маркер В клеток.
Ig A - Это первая линия противомикробной защиты. Преобладает в секретах желез (пот, слизь, слезы, молоко, вагинальный и простатический секреты).
Ig Е - Активирует тучные клетки и базофилы в аллергических реакциях (вызывает выделение гистамина, что способствует развитию отека). Не связывает комплемент.
Ig D - Маркер В ^, опосредует дифференцировку В клеток. Не связывает комплемент.
Антигены гистосовместимости — гликопротеины, существующие на поверхности всех клеток, получивших название молекул гистосовместимости (т.е. совместимости тканей).
Главные молекулы гистосовместимости — семейство гликопротеинов, кодируемое генами, составляющими главный комплекс гистосовместимости (МНС)
Существует два основных класса молекул МНС, каждый из которых представляет собой набор гликопротеинов клеточной поверхности. Молекулы МНС
класса I экспрессируются практически на всех клетках,
класса II — на клетках, участвующих в иммунных ответах (лимфоцитах, макрофагах). Молекулы класса I узнаются цитотоксическими Т-клетками, которые должны взаимодействовать с любой клеткой организма, оказавшейся зараженной вирусом, тогда как молекулы класса II узнаются Т-хелперами (Тх), которые взаимодействуют в основном с другими клетками, участвующими в иммунных ответах, такими как В-лимфоциты и макрофаги (антигенпредставляющие клетки).
Согласно клонально-селекционной теории иммунитета, в организме существуют многочисленные группы (клоны) лимфоцитов, генетически запрограммированные реагировать на один или несколько антигенов. При первой встрече с антигеном (первичный ответ) лимфоциты стимулируются и подвергаются трансформации в бластные формы, которые способны к пролиферации и дифференцировке в иммуноциты. В результате пролиферации увеличивается число лимфоцитов соответствующего клона, «узнавших» антиген. Дифференцировка приводит к появлению двух типов клеток — эффекторных и клеток памяти.
Эффекторные клетки непосредственно участвуют в ликвидации или обезвреживании чужеродного материала. К эффекторным клеткам относятся активированные лимфоциты и плазматические клетки.
Клетки памяти — это лимфоциты, возвращающиеся в неактивное состояние, но несущие информацию (память) о встрече с конкретным антигеном. При повторном введении данного антигена они способны обеспечивать быстрый иммунный ответ большей интенсивности (вторичный ответ) вследствие усиленной пролиферации лимфоцитов и образования иммуноцитов.
Различают дифференцировку и специализацию В- и Т-лимфоцитов
· антиген не зависимую
· антигензависимую.
Антигеннезависимая пролиферация и дифференцировка генетически запрограммированы на образование клеток, способных давать специфический тип иммунного ответа при встрече с конкретным антигеном благодаря появлению на плазмолемме лимфоцитов особых «рецепторов». Она совершается в центральных органах иммунитета (тимус, костный мозг или фабрициева сумка у птиц) под влиянием специфических факторов, вырабатываемых клетками, формирующими микроокружение (ретикулярная строма или ретикулоэпителиальные клетки в тимусе).
Антигензависимая пролиферация и дифференцировка Т- и В- лимфоцитов происходят при встрече с антигенами в периферических лимфоидных органах, при этом образуются эффекторные клетки и клетки памяти, сохраняющие информацию о действовавшем антигене. Образующиеся
· Т-л составляют пул долгоживущих, рециркулирующих лимфоцитов, а
· В-л - короткоживущих клеток.
Гуморальный иммунитет обеспечивается кооперативным взаимодействием клеток. Попавший в организма антиген поглощается макрофагом. Макрофаг расщепляет его на фрагменты, которые в комплексе в молекулами МНС класса II появляются на поверхности клетки. Такая обработка антигена макрофагом называется процессированием антигена.
Для дальнейшего развития иммунного ответа на антиген необходимо участие Т-х. Но прежде Т-х должны быть активированы сами. Эта активация происходит тогда, когда антиген обработанный макрофагом, распознается Т-х. Узнавание Т-х комплекса антиген + молекула МНС II класса на поверхности макрофага стимулирует секрецию интерлейкина I (ИЛ-I) макрофагом. Под воздействием ИЛ-I активизируются синтез и секреция ИЛ-II Т-х клеткой. Выделение Т-х клеткой ИЛ-II стимулирует ее пролиферацию. Т-х активируют В-л путем секреции ИЛ-2
Активация В-л происходит также при прямом взаимодействии антигена с иммуноглобулиновым рецептором В-клетки. В-лимфоцит сам процессирует антиген и представляет его фрагмент в комплексе с молекулой МНС II класса на клеточной поверхности. Этот комплекс узнает уже задействованный в иммунной реакции Т-х. Узнавание рецептором Тх-клетки комплекса «АГ+молекула МНС II класса» на поверхности В-лимфоцита приводит к секреции Тх интерлейкинов ИЛ-2, ИЛ-4, ИЛ-5, ИЛ - 6, у - ИФН (у -интерферона), под действием которых Вл-клетка размножается и дифференцируется с образованием плазматических клеток и В-клеток памяти.
В результате дедифференцировки образуются
Вб-л, при этом увеличивается ядро и цитоплазма клетки. Она вступает в активное состояние и превращается в
плазмобласт - крупная клетка с большим содержанием рибосом и слаборазвитой ЭПС. Ядро с эухроматином 1-2 ядрышка. Затем образуется
проплазмоцит - клетка становится меньше, в ней развивается ЭПС. Ядро расположено эксцентрично, хроматин в виде спиц колеса, имеется светлый дворик.
плазмоцит заполняется большим количеством расширенных цистерн гр-ЭПС, заполненная иммуноглобулинами. Окончательное формирование антител происходит в К. Гольджи путем присоединения полисахаридов. Антитела поступают в кровь и взаимодействуют с антигенами, образуя комплекс антиген + антитело, которое в последующем выпадает в осадок, а затем фагоцитируется.
Тс - супрессоры вместе с особой популяцией В-лимфоцитов подавляют выработку антител плазмоцитами.
Макрофаги участвуют как в естественном, так и в приобретенном иммунитете. Участие в естественном иммунитете проявляется в их способности к фагоцитозу, в синтезе фагоцитина, лизоцима, интерферона, пирогена, компонентов комплемента, контролируют размножение опухолевых клеток.
Их роль в приобретенном иммунитете заключается в пассивной передаче антигена Т- и В- лимфоцитам, в индукции специфического ответа на антигены. Макрофаги принимают участие в 2 стадиях иммунного ответа продуктивной и индуктивной:
1ст. индуктивная - макрофаги вырабатывают медиаторы, вызывающие пролиферацию лимфоцитов. 2ст. –продуктивная - когда они участвуют в выработке антител и разрушении антигена.
Участие тучных клеток и эозинофилов в иммунных реакциях. При первичном и особенно при повторном введении антигенов наблюдается увеличение тучных клеток, их контакт с макрофагами и массовая дегрануляция. Тучные клетки выделяют гепарин, гистамин и серотонин, оказывающие неспецифическое влияние на пролиферацию и дифференцировку иммунокомпетентных клеток (Т- и В-лимфоцитов).
Избыток гистамина приводит к увеличению числа эозинофилов, которые участвуют в его разрушении. В индуктивной фазе эозинофилы принимают участие в активизации макрофагов. В продуктивной фазе они выполняют дезинтоксикационную функцию, участвуя в фагоцитозе и разрушении комплекса антиген + антитело.

