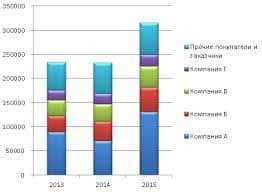
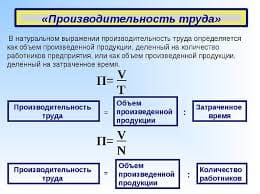
2017-12-16
2017-12-16 570
570Нарушение функций мозжечка. Нарушение функций мозжечка проявляется в различных видах нарушения движений.
1. Асинергия – отдельные компоненты двигательной программы выполняются не одновременно, а скорее последовательно (распад движения). Асинергия приводит к следующим феноменам:
1.1. Дисметрия – движения выполняются в избыточном или недостаточном объеме, а затем наступает их чрезмерная компенсация.
1.2. Церебеллярная атаксия – у таких больных наблюдается неверная походка с широко расставленными ногами и избыточными движениями.
2. Интенционный тремор – тремор, который отсутствует в покое, но возникает при движении.
3. Гипотония – снижение мышечного тонуса. Этот симптом часто сопровождается слабостью и быстрой утомляемостью мышц (связано главным образом с повреждениями полушарий мозжечка).
4. Нистагм глаз.
5. Головокружение.
6. Дефекты речи.
Типичным проявлением поражений мозжечка служит триада Шарко: нистагм, интенционный тремор и скандированная речь.
Вопрос 51. Физиологические механизмы образования условных рефлексов, их структурно-функциональная основа. Развитие преставлений И.П. Павлова о механизмах формирования временных связей.
Временная (условная) связь — это совокупность нейрофизиологических, биохимических и ультраструктурных изменений мозга, возникающих в процессе сочетания условного и безусловного раздражителей и формирующих определенные взаимоотношения между различными мозговыми образованиями. Механизм памяти фиксирует эти взаимоотношения, обеспечивая их удержание и воспроизведение. В зоне коркового представительства условного стимула и коркового (или подкоркового) представительства безусловного стимула формируются два очага возбуждения. Очаг возбуждения, вызванный безусловным стимулом внешней или внутренней среды организма, как более сильный (доминантный) притягивает к себе возбуждение из очага более слабого возбуждения, вызванного условным стимулом. После нескольких повторных предъявлений условного и безусловного раздражителей между этими двумя зонами «проторяется» устойчивый путь движения возбуждения: от очага, вызванного условным стимулом, к очагу, вызванному безусловным стимулом. В результате изолированное предъявление только условного стимула теперь приводит к реакции, вызываемой ранее безусловным стимулом.В качестве главных клеточных элементов центрального механизма образования условного рефлекса выступают вставочные и ассоциативные нейроны коры большого мозга. Корковый пункт условного раздражителя в принципе имеет такую же структуру, как корковое представительство безусловного рефлекса, так как условный раздражитель сам по себе вызывает соответствующую безусловно-рефлекторную реакцию, особенно, если в качестве условного раздражителя взять какой-нибудь механический, электрический или химический раздражитель. Поэтому при сочетании условного и безусловного раздражителей временная связь по существу замыкается между корковыми представительствами двух безусловных рефлексов, т.е. между нейронами корковых ветвей их рефлекторных дуг. Следовательно, процесс образования условного рефлекса можно рассматривать как корковый синтез двух (или нескольких) безусловных рефлексов. Такое представление о месте и характере замыкания временной связи подтверждается экспериментальными исследованиями Э.А. Асратяна. Если подобрать два безусловных раздражителя примерно одинаково физиологической силы, например, пищевой и оборонительный, а затем сочетать их строго попеременно (то в одной, то в обратной последовательности), то образуется условный рефлекс в необычной форме. Каждый из безусловных раздражителей будет вызывать как двигательную, так и секреторную реакцию. Электрокожное раздражение становится сигналом пищи, а пища - сигналом электрокожного раздражения. Следовательно‑ временные связи могут быть двусторонними.
Физиологические механизмы замыкания временной связи. Образование временной связи, по Павлову, является результатом взаимодействия двух одновременно возбуждаемых пунктов коры. Наличие двух очагов возбуждения в коре закономерно вызовет движение процесса возбуждения от более слабого (вызванного индифферентным раздражителем) к более сильному (вызванному безусловным раздражителем). Таким образом, в основе механизма замыкания временной связи Павлов усматривал явление типа проторения пути, суммационного рефлекса, доминанты.
Опыты Русинова показывают, что если действием постоянного тока на участок коры мозга вызвать в нем повышенную возбудимость, то он приобретает свойства доминантного и любое раздражение прежде всего вызывает теперь реакцию, связанную с возбуждением такого очага. Если это двигательный центр, то звук вызывает движение лапы. Искусственно созданный доминантный очаг сохраняется некоторое время и после выключения тока. Однако обычно доминантный очаг сохраняется сравнительно недолго, тогда как образовавшаяся временная связь является стойкой. Предполагают поэтому, что доминантный механизм играет роль лишь в первой стадии образования условного рефлекса, в процессе прокладывания временной связи, т.е. в образовании проходимости ранее бездействовавших синапсов вставочных нейронов. Упрочнение же временной связи осуществляется по другом механизму. Эти механизмы пока точно не изучены. Одни исследователи главным считают функциональную перестройку синапсов под влиянием повторных раздражений. Другие считают, что все дело в соответствующем изменении лабильности клеток коры в центрах рефлексов, третьи предполагают, что поддержание стойкого состояния постоянной проводимости импульсов в образовавшихся временных связях обусловлено движением импульсов по кольцевым системам коры.
Беритов и Ройтбак основными в процессе замыкания временной связи считали морфологические перестройки в ЦНС под влиянием сочетаний (утолщение нейрофибрилл, миэлинизация нервных пресинаптических волокон, появление новых синапсов и т.п.).
Клеточные механизмы замыкания временной связи. В основе механизмов формирования ассоциативной связи лежит принцип конвергентного замыкания. Идея о конвергенции гетерогенных стимулов как принципе установления условного рефлекса была выдвинута Дж. Экклзом (1968). Он предположил существование центра конвергенции, который не имеет определенной локализации. На нейронах, входящих в состав этого центра, встречаются условный и безусловный сигналы. При этом происходит формирование пространственно-временной структуры возбуждения, которая затем передается в нейроны нижеследующего уровня.
Сохранив принцип конвергенции стимулов гетерогенных модальностей как исходный механизм взаимодействия и интеграции, П.К. Анохин (1968, 1974) дополнил концепцию представление о том, что взаимодействие сигналов различных сенсорных и биологических модальностей происходит на одном нейроне (рис. 4). Особые свойства подкрепляющего фактора определяются специфическими химическими реакциями, которые он вызывает, и соответствующими молекулярными преобразованиями в пределах мембраны и цитоплазмы нейрона. Здесь происходит ассоциация эффектов условной и безусловной стимуляции с последующими выходами сложившейся интеграции на аксон в виде клеточного разряда. Данная теоретическая концепция является основой для проведения исследований механизмов формирования ассоциативного обучения на одном нейроне.
Микроэлектродные исследования показали, что формирование ассоциативных связей происходит на всех уровнях мозга млекопитающих. Исследованы нейроны коры, ретикулярной формации, лимбических структур, стриопаллидарной системы, гипоталамуса. Показано, что до 60% нейронов различных корковых и подкорковых структур мозга способны формировать условные ответы на ранее неэффективные стимулы.
Внутриклеточные механизмы ассоциативного обучения. Опыты с ассоциативным обучением изолированных нейронов показали, что около 15% из них обучаются в процессе предъявления сочетаний. и около 80% - демонстрировали феномен отсроченного обучения, когда реакция на условный сигнал появлялась не сразу, а через какое-то время. Независимо от вида ассоциируемых стимулов и от особенностей предъявления сочетаний, каждый конкретный нейрон всегда обучается по одному и тому же способу – или во время обучения, или отсроченно. Это качество является его индивидуальной характеристикой. В основе этого феномена лежат особенности внутриклеточных процессов, опосредующих ассоциативное обучение, и эти процессы различны по скорости их развития.
Эффективность входа в нейрон может меняться под влиянием разной степени эффективности последующих возбуждений, поступающих по другим входам в этот же нейрон. С этой точки зрения особый интерес представляют командные нейроны, рецептивное поле которых состоит из широкой пластичной зоны и узкой области высоко стабильных реакций. При повторении стимула на пластичную зону рецептивного поля реакция угасает. При нанесении стимула на зону стабильных реакций ответы нейрона устойчиво сохраняются. При комбинации раздражений малоэффективной зоны с раздражениями области эффекторных реакций ответы со слабо эффективной зоны усиливаются. В пределах одного нейрона неэффективный вход может стать эффективным при сочетании его возбуждения с возбуждением высокоэффективного входа. С этой точки зрения формирование временной связи возможно в отдельном нейроне.