 2018-01-08
2018-01-08 1716
1716Введение
Одним из основных достижений биологии конца XX века явилось выявление трех эволюционных ветвей жизни, сформировавшихся на заре ее возникновения, - это домены бактерий, архей и эукариотов. Археи, как полагают, являются одними из самых древних организмов на Земле, и до последнего времени вопрос о точном времени их происхождения оставался открытым. На сегодняшний день самая старая находка датируется возрастом 2,71 - 2,65 миллиарда лет. Археи, несомненно, сыгравшие существенную роль в эволюции биосферы, и в наше время занимают в ней своеобразное место, обеспечивая прохождение определенных процессов круговорота веществ, являющегося необходимым условием ее существования. Археи чрезвычайно распространены в мире, но известно о них меньше, чем о бактериях. Характерные черты, заметные лишь учёным, делают археев абсолютно неповторимыми, чётко отличая их от всех прочих - бактерий, грибов, растений и животных. Вероятно, многие из этих микроорганизмов состоят из химических структур, неизвестных науке. Археи имеют важное промышленное значение, так как только они могут образовывать метан.
Архе́и (лат. Archaea от др.-греч. ἀρχαῖος «извечный, древний, первозданный, старый») — домен живых организмов (по трёхдоменной системе Карла Вёзе наряду с бактериями и эукариотами). Археи представляют собой одноклеточные микроорганизмы, не имеющие ядра, а также каких-либо мембранных органелл. Ранее археи объединяли с бактериями в общую группу, называемую прокариоты (или царство Дробянки (лат. Monera)), и они назывались архебактерии, однако сейчас такая классификация считается устаревшей[1]: установлено, что археи имеют свою независимую эволюционную историю и характеризуются многими биохимическими особенностями, отличающими их от других форм жизни. В настоящее время археи подразделяют на более чем 5 типов. Из этих групп наиболее изученными являются кренархеоты (лат. Crenarchaeota) и эвриархеоты (лат. Euryarchaeota). Классифицировать археи по-прежнему сложно, так как подавляющее большинство из них никогда не выращивались в лабораторных условиях и были идентифицированы только по анализу нуклеиновых кислот из проб, полученных из мест их обитания. Археи и бактерии очень похожи по размеру и форме клеток, хотя некоторые археи имеют довольно необычную форму, например, клетки Haloquadratum walsbyi плоские и квадратные. Несмотря на внешнее сходство с бактериями, некоторые гены и метаболические пути архей сближают их с эукариотами (в частности ферменты, катализирующие процессы транскрипции и трансляции). Другие аспекты биохимии архей являются уникальными, к примеру, присутствие в клеточных мембранах липидов, содержащих простую эфирную связь. Большая часть архей — хемоавтотрофы. Они используют значительно больше источников энергии, чем эукариоты: начиная от обыкновенных органических соединений, таких как сахара, и заканчивая аммиаком, ионами металлов и даже водородом. Солеустойчивые археи — галоархеи (лат. Haloarchaea) — используют в качестве источника энергии солнечный свет, другие виды архей фиксируют углерод, однако, в отличие от растений и цианобактерий, ни один вид архей не делает и то, и другое одновременно. Размножение у архей бесполое: бинарное деление, фрагментация и почкование. В отличие от бактерий и эукариот, ни один известный вид архей не формирует спор. Изначально архей считали экстремофилами, живущими в суровых условиях — горячих источниках, солёных озёрах, однако потом они были обнаружены в самых различных местах включая почву, океаны, болота и толстую кишку человека. Архей особенно много в океанах, и, возможно, планктонные археи являются самой многочисленной группой ныне живущих организмов. В наше время археи признаны важной составляющей жизни на Земле. Они играют роль в круговоротах углерода и азота. Ни один из известных представителей архей не является ни паразитом, ни патогенным организмом, однако они часто бывают мутуалистами и комменсалами. Некоторые представители являются метаногенами и обитают в пищеварительном тракте человека и жвачных, где они помогают осуществлять пищеварение. Метаногены используются в производстве биогаза и при очистке канализационных сточных вод, а ферменты экстремофильных микроорганизмов, сохраняющие активность при высоких температурах и в контакте с органическими растворителями, находят своё применение в биотехнологии.
Рис 1
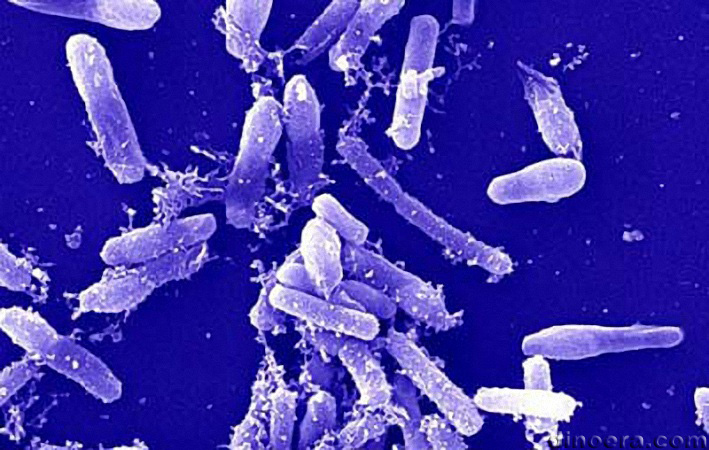
Рис 2
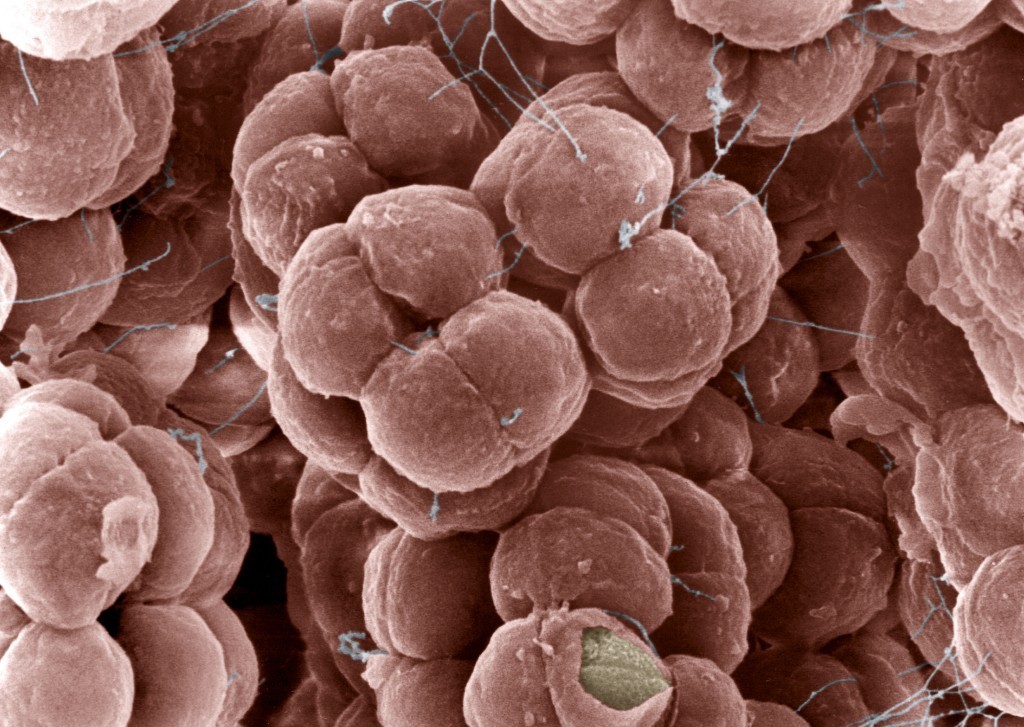
Морфология архей
Средний диаметр клеток архей 1 мкм, типичный для большинства известных прокариот. Клетки Nanoarchaeum equitans – самые маленькие среди архей, диаметром до 0,4 мкм. Помимо обычных для прокариот форм клеток в виде кокков (в том числе, с выступами или с дольками) или палочек (прямых или спиралевидных), среди архей также найдены клетки в виде почтовых марок и треугольников. Жгутики архей могут располагаться на 1–2 полюсах клеток или по всей поверхности. Поскольку белок флагеллин, составляющий основу жгутиков эубактерий, неустойчив в кислой среде, то у архей он замещается гликопротеинами, из которых состоят бактериальные пили. Собственно пили у архей не описаны. В отличие от других микроорганизмов, археи в составе мембранных липидов имеют не жирные кислоты, а многоатомные спирты, обычно с 20 или 40 атомами углерода. В последнем случае липидная пластина мембраны образована мономолекулярным слоем, что придает мембране особую прочность. У архей отсутствует пептидогликановая клеточная стенка, поэтому они устойчивы к пенициллину. Над плазматической мембраной они имеют слой белков или гликопротеинов. Грамположительные археи характеризуются наличием псевдомуреина в стенках, в состав которого помимо N-ацетилглюкозамина, присутствующего и в эубактериальных муреиновых клеточных стенках, входит ранее неизвестная N-ацетилталозаминуроновая кислота. Однако при окрашивании псевдомуреин часто разрушается. Поэтому лишь некоторые метаногены (в частности, Methanobacterium formicicum и терминальные пробки Methanospirillum hungatei) имеют грамположительную окраску Мембраны архей отличаются от мембран клеток из других доминионов по 3 основным признакам: 1. L-изомер глицерина – вместо D-глицерина в мембранах бактерий и эукариот; 2. Насыщенный полиизопрен – фитанил (3,7,11,15 тетраметилгексадесил) с 4 боковыми СH3-группами вместо неразветвлённых жирных кислот длиной 16–18 атомов углерода для ранее известных мембран; 3. Простые эфирные связи между L-глицерином и фитанилом, имеющие лишь один атом кислорода. В отличие от бактерий в мембранах архей и эукариот присутствует фосфатидилинозит. Как правило, археи имеют одиночную кольцевую хромосому, размер которой может достигать 5 751 492 пар нуклеотидов у Methanosarcina acetivorans, обладающей самым большим известным геномом среди архей. Одну десятую размера этого генома составляет геном с 490 885 парами нуклеотидов у Nanoarchaeum equitans, имеющего самый маленький известный геном среди архей; он содержит лишь 537 генов, кодирующих белки. Также у архей обнаружены более мелкие независимые молекулы ДНК, так называемые плазмиды. Возможно, плазмиды могут передаваться между клетками при физическом контакте, в ходе процесса сходного с конъюгацией бактерий. Археи могут поражаться вирусами, содержащими двухцепочечную ДНК. Эти виды неродственны другим группам вирусов и имеют различные необычные формы, включая бутылки, крючки и капли. Эти вирусы были тщательно изучены на термофилах, в основном отрядов Sulfolobales и Thermoproteales. В 2009 году был открыт вирус, содержащий одноцепочечную ДНК и поражающий галофильные археи. Защитные реакции архей против вирусов могут включать механизм, близкий к РНК-интерференции эукариот. Археи генетически отличны от эукариот и бактерий, причём до 15 % белков, кодируемых одним геномом археи, уникальны для этого домена, хотя функции большинства этих белков неизвестны. Большая часть уникальных белков, функция которых известна, принадлежит эвриархеотам и задействована в метаногенезе. Белки, общие для архей, бактерий и эукариот, участвуют в основных клеточных функциях и касаются в основном транскрипции, трансляции и метаболизма нуклеотидов. К другим особенностям архей можно отнести организацию генов, выполняющих связанные функции (к примеру, гены, ответственные за разные этапы одного и того же метаболического процесса), в опероны и большие отличия в строении генов тРНК и их аминоацил-тРНК синтетаз. Археи размножаются бесполым путём: бинарным или множественным делением, фрагментацией или почкованием. Клеточное деление определяется клеточным циклом: после того, как хромосома реплицировалась и две дочерние хромосомы разошлись, клетка делится. Детали были изучены лишь у рода Sulfolobus, но особенности его цикла очень схожи с таковыми и у эукариот, и у бактерий. Репликация хромосом начинается с множественных точек начала репликации с помощью ДНК-полимеразы, похожей на аналогичные ферменты эукариот. Однако белки, управляющие клеточным делением, такие как FtsZ, которые формируют сжимающее кольцо вокруг клетки, и компоненты септы, проходящей через центр клетки, схожи с их бактериальными эквивалентами. Археи не образуют споры. Некоторые виды Haloarchaea могут претерпевать смену фенотипа и существовать как клетки нескольких различных типов, включая толстостенные клетки, устойчивые к осмотическому шоку и позволяющие археям выживать в воде с низкой концентрацией соли. Однако эти структуры не служат для размножения, а скорее помогают археям осваивать новые среды обитания. Клетки некоторых видов архей могут объединяться в агрегаты и филаменты длиной до 200 мкм. Эти организмы могут формировать биоплёнки. В культурах Thermococcus coalescens клетки сливаются друг с другом, формируя одну крупную клетку. Археи рода Pyrodictium образуют сложные многоклеточные колонии, в которых клетки объединены с помощью длинных, тонких, полых трубок, называемых cannulae, которые выступают над поверхностями клеток и собирают их в густое кустовидное скопление. Функции этих трубок не ясны, но, возможно, они осуществляют коммуникацию и обмен питательными веществами между соседними клетками.
| Свойственно археям и бактериям | Свойственно археям и эукариотам | Свойственно только археям |
| Нет оформленного ядра и мембранных органелл | Нет пептидогликана (муреина) | Структура клеточной стенки (к примеру, клеточные стенки некоторых архей содержат псевдомуреин) |
| Кольцевая хромосома | ДНК связана с гистонами | В клеточной мембране присутствуют липиды, содержащие простую эфирную связь |
| Гены объединены в опероны | Трансляция начинается с метионина | Структура флагеллинов |
| Схожие РНК-полимераза, промоторы и другие компоненты транскрипционного комплекса, есть интроны и процессинг РНК | Структура рибосом (некоторые признаки сближают с бактериями, некоторые — с эукариотами) | |
| Полицистронная мРНК | Схожие репликация и репарация ДНК | Структура и метаболизм тРНК |
| Размер клеток (более чем в 100 раз меньше, чем у эукариот) | Схожая АТФаза (тип V) | Нет синтазы жирных кислот |
Сравнительная таблица архей и других доменов
Экология архей
Археи распространены повсеместно. Начиная от гидротермальных источников, кратеров вулканов и дна Северного Ледовитого океана и заканчивая столь уютным для них пищеварительным трактом людей и термитов. За счет различных путей метаболизма, археи являются важными членами многих экологических сообществ. Наиболее интересными с точки зрения метаболизма являются метаногенные археи, которые, как ни странно, являются единственным биологическим источником метана на земле. Считаются, что именно такой путь метаболизма использовался первыми организмами на Земле. Тесно взаимодействуя с разнообразием биологических видов, они совместно окисляют органику в анаэробных условиях. Важно, что синтрофическое питание необходимо для каждого из членов полимикробного сообщества, ведь в одиночку каждый вид не может использовать субстраты для роста ввиду термодинамических препятствий. Дело в летучих жирных кислотах (volatile fatty acids — VFA) – ацетат, пропионат, бутират, которые являются нежелательными для анаэробов, поскольку их окислительный метаболизм до H2 и CO2возможен только при низком парциальном давлении H2 вне клетки. Именно за счет утилизации H2 и CO2 метаногенами в реакциях синтеза метана и поддерживается оптимальное давление водорода, что поддерживает метаболизм анаэробных бактерий на высоком уровне. Этим механизмом достигается сохранение энергии в сообществах и на нем же основан межвидовой транспорт водорода. Соответственно, антагонистические отношения складываются у Архей с другими потребителями H2 (в частности – Treponema spp., сульфат- и сероредуцирующие бактерии)
Таблица:типы питания архей
| Тип питания | Источник энергии | Источник углерода | Примеры |
| Фототрофы | Свет | Органические соединения | Halobacterium (у них имеется галородопсин) |
| Литотрофы | Неорганические соединения (сера, аммиак) | Органические соединения или фиксация углерода | Ferroglobus, Methanobacteria, Pyrolobus |
| Органотрофы | Органические соединения | Органические соединения или фиксация углерода | Pyrococcus, Sulfolobus, Methanosarcinales |
Взаимодействие с другими организмами
Метанообразующие археи вступают в симбиоз с термитами Хорошо изученные отношения между археями и другими организмами — мутуализм и комменсализм. Пока не существует чётких доказательств существования патогенных или паразитических видов архей. Однако была предположена связь между некоторыми видами метаногенов и инфекциями полости рта. Кроме того, вид Nanoarchaeum equitans, возможно, является паразитом другого вида архей, поскольку он выживает и размножается только в клетках кренархеона Ignicoccus hospitalis и не приносит никакой очевидной выгоды своему хозяину. С другой стороны, архееподобные ацидофильные наноорганизмы Ричмондских рудников (англ. Archaeal Richmond Mine acidophilic nanoorganisms) (ARMAN) иногда прикрепляются к клеткам других архей в биоплёнках кислых сточных вод рудников. Природа этого взаимодействия не ясна, но, в отличие от случая Nanorchaeaum—Ignicoccus, сверхмелкие клетки ARMAN всегда остаются независимыми от клеток Thermoplasmatales.
Мутуализм
Одним хорошо понятным примером мутуализма является взаимодействие простейших и метанообразующих архей, обитающих в пищеварительном тракте животных, способных переваривать целлюлозу, таких как жвачные и термиты. В этих анаэробных условиях простейшие разлагают целлюлозу для получения энергии. В этом процессе в качестве побочного продукта освобождается водород, однако высокий его уровень сокращает получение энергии. Метаногены превращают водород в метан, и простейшие могут дальше нормально получать энергию. В случае анаэробных простейших вроде Plagiopyla frontata археи живут внутри клетки простейшего и потребляют водород, образуемый в его гидрогеносомах. Археи также взаимодействуют и с более крупными организмами. Например, морская архея Cenarchaeum symbiosum живёт внутри (как эндосимбионт) губки Axinella mexicana.
Комменсализм
Археи могут быть комменсалами, то есть существовать совместно с другим организмом, не принося ему ни пользы, ни вреда, но с выгодой для себя. К примеру, метаноген Methanobrevibacter smithii является наиболее типичным представителем архей в микрофлоре человека. Каждый десятый прокариот в человеческом пищеварительном тракте принадлежит к этому виду. В пищеварительном тракте термитов и человека эти метаногены в действительности могут быть мутуалистами, взаимодействущими с другими микробами пищеварительного тракта и способствующими пищеварению. Археи также взаимодействуют с другими организмами, к примеру, живут на внешней поверхности кораллов и в части почвы, прилегающей к корням растений (ризосфере)
Типы архей
· Crenarchaeota (Креноархеоты) — термофилы, термоацидофилы, серные анаэробные бактерии;
· Euryarchaeota (Эвриархеоты) — метаногенные и галофильные археи;
· Nanoarchaeota (Наноархеоты) — единственный известный представитель Nanoarchaeum equitans;
· Korarchaeota (Корархеоты)— ДНК обнаружена в геотермальных источниках США, Исландии, на рисовых полях Японии, культивируемые виды пока неизвестны;
· Thaumarchaeota (Таумархеоты)— в основном окислители аммония, как например, морской аммоний-окислитель Nitrosopumilus maritimus и аммоний-окислитель преимущественно почвенного происхождения Nitrososphaera gargensis. Недавние филогенетические исследования, основанные на сравнительном анализе структур рибосомальных белков и других важнейших генов, подтвердили существование типа Таумархеот. Был выделен в 2008 году
· Иногда выделяют также шестой тип — Aigarchaeota (Аигархеоты) к которому относится единственный вид Caldiarchaeum subterraneum.
В мае 2015 года были опубликованы результаты исследования, предлагающего выделение нового типа архей — Lokiarchaeota (Локиархеоты) с предполагаемым родом Lokiarchaeum. Он был выделен на основании генома, собранного при метагеномном анализе образцов, полученных рядом с гидротермальными источниками в Атлантическом океане на глубине 2,35 км. Филогенетический анализ показал, что Lokiarchaeota и эукариоты образуют монофилетический таксон. Геном Lokiarchaeota содержит около 5400 генов, кодирующих белки. Среди них были обнаружены гены, близкие к генам эукариот. В частности, гены, кодирующие белки, отвечающие за изменение формы клеточной мембраны, определение формы клетки и динамический цитоскелет. Результаты этого исследования служат подтверждением так называемой двухдоменной, или эоцитной гипотезы, согласно которой эукариоты появились как особая группа внутри архей, близкая к Lokiarchaeota и приобретшая митохондрии в результате эндосимбиоза,. В январе 2016 года были опубликованы результаты метагеномных исследований по реконструкции геномов архей из морских осадочных отложений, которые свидетельствуют об обнаружении нового типа архей — Thorarchaeota (Торархеоты). Организмы этой группы способны к образованию ацетата при деградации белков. Они также имеют гены, необходимые для восстановления элементарной серы и тиосульфата, поэтому эти организмы участвуют в круговороте серы В итоге к настоящему времени известны 8 типов архей. Однако хорошо изучено лишь 2 из них: кренархеоты и эвриархеоты








