 2018-01-08
2018-01-08 6171
6171Вопросы для подготовки
- Учение И.П.Павлова об анализаторных системах. Строение анализаторной системы, взаимосвязь отделов анализатора между собой. Значение анализаторов.
- Роль анализаторов в формировании высшей нервной деятельности.
- Морфофункциональная характеристика зрительного анализатора. Оптическая система глаза. Зрачковый и аккомодационный рефлексы.
- Рецепторный отдел зрительного анализатора. Сетчатка глаза, физиология свето- и цветовосприятия. Проводниковый и корковый отделы зрительного анализатора.
- Физиология слухового анализатора. Характеристика процессов звукопроведения, рецепция звука. Проводниковый и корковый отделы слухового анализатора.
- Физиология кожного анализатора. Виды кожных рецепторов. Проводниковый и корковый отделы.
- Физиология интероцептивного анализатора. Роль этого анализатора в поддержании гомеостаза. Особенности анализатора.
- Современные представления о ноцицепции и антиноцицепции. Нейрохимические механизмы. Понятие об антиноцицепции.
- Двигательный анализатор. Значение афферентации от проприорецепторов и рецепторов вестибулярного аппарата.
ДОМАШНЕЕ ЗАДАНИЕ
|
|
|
1. Дайте определение понятия анализаторная система.
Анализаторные системы- это сложные нервные структуры, начинающиеся переферическими воспринимающимися образованиями (рецепторами) и заканчивающимися нервными центрами, расположенными в высших отделов мозга и обеспечивающих анализ воспринятых раздражений и выработку на этой основе сигналов для построения ответной деятельности организма.
2. Укажите функции рецепторных частей сенсорных систем.
Сенсорная система выполняет следующие основные функции, или операции, с сигналами: 1) обнаружение; 2) различение; 3) передачу и преобразование; 4) кодирование; 5) детектирование признаков; 6) опознание образов. Обнаружение и первичное различение сигналов обеспечивается рецепторами, а детектирование и опознание сигналов — нейронами коры больших полушарий. Передачу, преобразование и кодирование сигналов осуществляют нейроны всех слоев сенсорных систем.
3. Рецепторный и генераторный потенциал. Особенности передачи информации в первично и вторично чувствующих системах.
Общие механизмы возбуждения рецепторов. При действии стимула на рецепторную клетку происходит преобразование энергии внешнего раздражения в рецепторный сигнал, или трансдукция сенсорного сигнала.
Этот процесс включает в себя три основных этапа:
1) взаимодействие стимула, т. е. молекулы пахучего или вкусового вещества (обоняние, вкус), кванта света (зрение) или механической силы (слух, осязание) с рецепторной белковой молекулой, которая находится в составе клеточной мембраны рецепторной клетки;
|
|
|
2) внутриклеточные процессы усиления и передачи сенсорного стимула в пределах рецепторной клетки;
3) открывание находящихся в мембране рецептора ионных каналов, через которые начинает течь ионный ток, что, как правило, приводит к деполяризации клеточной мембраны рецепторной клетки (возникновению так называемого рецепторного потенциала).
В первично-чувствующих рецепторах этот потенциал действует на наиболее чувствительные участки мембраны, способные генерировать потенциалы действия — электрические нервные импульсы. Во вторично-чувствующих рецепторах рецепторный потенциал вызывает выделение квантов медиатора из пресинаптического окончания рецепторной клетки. Медиатор (например, ацетилхолин), воздействуя на постсинаптическую мембрану первого нейрона, изменяет ее поляризацию (генерируется постсинаптический потенциал). Постсинаптический потенциал первого нейрона сенсорной системы называют генераторным потенциалом, так как он вызывает генерацию импульсного ответа. В первично-чувствующих рецепторах рецепторный и генераторный потенциалы — одно и то же.
4. Функции проводниковой части сенсорных систем.
Проводниковый отдел анализатора – цепь нейронов, находящихся в разных слоях на каждом уровне ЦНС. Афферентные (периферические) и промежуточные нейроны стволовых и подкорковых структур ЦНС обеспечивает: проведение возбуждения от рецепторов в кору большого мозга и частичную переработку информации.
Проведение возбуждения осуществляется 2-мя афферентными путями:
1) Специфический проекционный путь (прямые афферентные пути) – от рецептора по строго обозначенным специфическим путям с переключением на различных уровнях ЦНС (на уровне СМ и продолговатого мозга, в зрительных буграх и в соответствующей проекционной зоне коры большого мозга).
2) Неспецифический путь – с участием РФ. На уровне ствола мозга от специфического пути отходят коллатерали к клеткам РФ, к которым могут конвергировать (сходиться) различные афферентные возбуждения, обеспечивая взаимодействие анализаторов. При этом афферентные возбуждения теряют свои специфические свойства (сенсорную модальность) и изменяют возбудимость корковых нейронов
Морфофункциональная организация корковых отделов анализаторных систем.
Кора большого мозга имеет следующие морфофункциональные особенности:
— многослойность расположения нейронов;
— модульный принцип организации;
— соматотопическая локализация рецептирующих систем;
— экранность, т. е. распределение внешней рецепции на плоскости нейронального поля коркового конца анализатора;
— зависимость уровня активности от влияния подкорковых структур и ретикулярной формации;
— наличие представительства всех функций нижележащих структур ЦНС;
— цитоархитектоническое распределение на поля;
— наличие в специфических проекционных сенсорных и моторной системах вторичных и третичных полей с ассоциативными функциями;
— наличие специализированных ассоциативных областей;
— динамическая локализация функций, выражающаяся в возможности компенсаций функций утраченных структур;
— перекрытие в коре большого мозга зон соседних периферических рецептивных полей;
— возможность длительного сохранения следов раздражения;
— реципрокная функциональная взаимосвязь возбудительных и тормозных состояний;
— способность к иррадиации возбуждения и торможения;
— наличие специфической электрической активности.
Виды фоторецепторов, механизмы фоторецепции.
К фоторецепторам в сетчатке глаза человека относятся 3 вида колбочек (каждый тип возбуждается светом определенной длины волны), которые отвечают за цветное зрение, и один вид палочек, который отвечает за сумеречное зрение. В сетчатке глаза человека насчитывается 110 ÷ 125 млн палочек и 4 ÷ 7 млн колбочек.
|
|
|
Таблица, иллюстрирующая различия между палочками и колбочками (по книге Эрика Канделя «Принципы науки о нейронах»[6])
| Палочки | Колбочки |
| Используются для ночного зрения (в условиях слабой освещенности) | Используются для дневного зрения (в условиях высокой освещенности) |
| Высокочувствительны; воспринимают и рассеянный свет | Не очень чувствительны к свету; реагируют только на прямой свет |
| Повреждение вызывает никталопию (гемералопию) | Повреждение вызывает слепоту, дневную слепоту, ахроматопсию |
| Низкая острота зрения | Высокая острота зрения; лучшее пространственное разрешение |
| Нет в центральной ямке | Сосредоточены в центральной ямке |
| Замедленная реакция на свет | Быстрая реакция на свет, могут воспринимать более быстрые изменения у раздражителя |
| Имеют больше пигмента, чем колбочки | Имеют меньше пигмента |
| Мембранные диски не привязаны непосредственно к клеточной мембране | Мембранные диски крепятся к наружной мембране |
| В 20 раз больше, чем колбочек, по количеству. | |
| Один тип фоточувствительного пигмента | Три типа фоточувствительных пигментов у человека |
| Ср. Ахроматическое зрение | Ср. Цветное зрение |
5. Перечислите оптические среды глаза.Укажите преломляющую силу роговицы, хрусталика, стекловидного тела.
1) Роговица - является наиболее мощным преломляющим элементом, по сути является собирающей линзой.
2) Водянистая влага передней камеры - является прозрачной и по сути не влияет на преломление, однако является оптической средой.
3) Хрусталик - имеет вид двояковыпуклой линзы, способен изменять свою кривизну. Благодаря хрусталику мы четко видим предметы на разном расстоянии от нас (аккомодация). Пройдя за хрусталик световые лучи медленно сходятся и переворачиваются ближе к заднему полюсу глаза.
4) Стекловидное тело - находится за хрусталиком, также является прозрачной и не влияет на преломление, однако причисляется к оптическим средам глаза.
|
|
|
6. Нарисуйте схему рефлекторного пути зрачкового и аккомодационного рефлексов. Укажите их физиологическое значение.
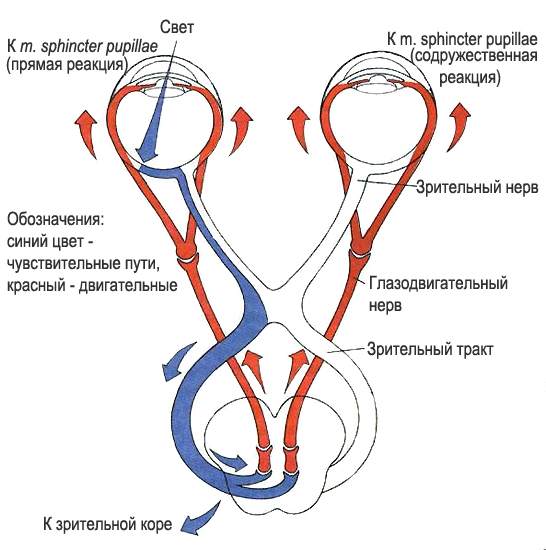
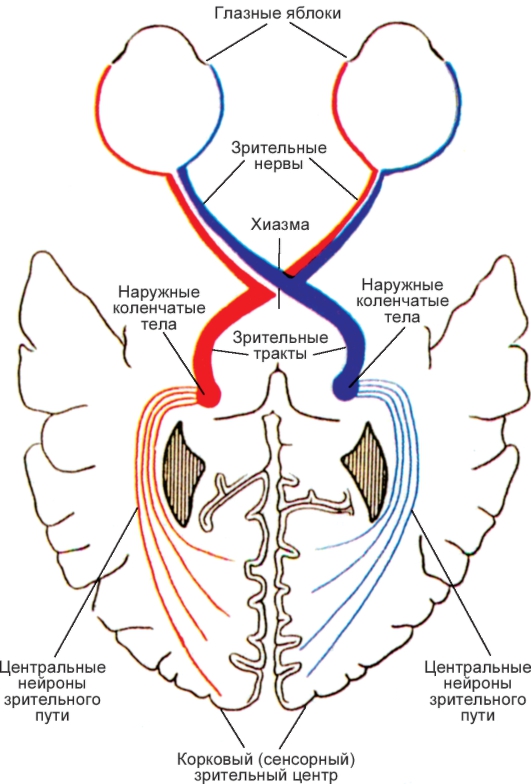
7. Дайте определение бинаурального слуха, опишите механизм его формирования и значение для восприятия звуков.
Бинауральный слух (от лат. bini -два + auricula - ухо) - восприятие звуков с помощью обоих ушей и симметричных (правой и левой) частей слуховой системы. Б. с. позволяет локализовать источник звука в пространстве за счет детекции различий основных характеристик звуковых сигналов, поступающих на разные уши. При этом наиболее точная дифференциальная локализация звуков наблюдается, когда интенсивность сигналов равна 70-100 дБ над порогом слышимости.
Природа не случайно предоставила человеку возможность слышать двумя ушами. Орган слуха человека содержит два уха и билатеральные невральные слуховые пути и структуры, которые предназначены не только для того, чтобы хорошо слышать, но и обнаруживать, различать и понимать звуки в различных, и особенно в сложных акустических условиях. При нормальном слухе акустические сигналы от правого и левого уха по восходящим нервным ипси- и контралатеральным слуховым путям поступают и обрабатываются соответствующими отделами головного мозга и поступают в билатеральную слуховую кору (рис).
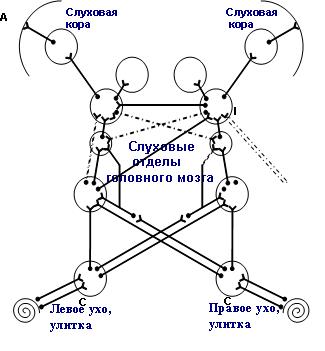 Билатеральное строение слухового органа дает возможность нормально слышащему человеку легко и уверенно локализовать источники звука и определять направление их движения в пространстве. Его способность понимать разговорную речь в сложных акустических условиях также обеспечивается структурами и механизмами бинаурального взаимодействия в центральных отделах слуховой системы. Для этого она использует различные приемы, такие как учет акустической тени головы, восприятие интерауральных (межушных) различий в воспринимаемых сигналах по интенсивности, времени и спектральному составу, включая механизмы бинауральной демаскировки и суммации звуков (1 - 4).
Билатеральное строение слухового органа дает возможность нормально слышащему человеку легко и уверенно локализовать источники звука и определять направление их движения в пространстве. Его способность понимать разговорную речь в сложных акустических условиях также обеспечивается структурами и механизмами бинаурального взаимодействия в центральных отделах слуховой системы. Для этого она использует различные приемы, такие как учет акустической тени головы, восприятие интерауральных (межушных) различий в воспринимаемых сигналах по интенсивности, времени и спектральному составу, включая механизмы бинауральной демаскировки и суммации звуков (1 - 4).
Бинауральные механизмы, такие как бинауральная демаскировка и суммация, обеспечивают слушателю улучшение соотношение сигнал / помеха от 3 до 6 дБ и более. Эти центральные механизмы бинаурального слуха являются важной физиологической базой для улучшения разборчивости речи при маскировке шумом и обеспечивают основные коммуникативные преимущества бинаурального слуха в шумных условиях (3-5).
Разборчивость и понимание звуков речи в шуме зависит от способности бинаурального слуха выделять и обрабатывать интерауральные или дирекционные признаки от каждого отдельного источника звука, в том числе речи и шума. В этой связи у пользователя с двумя слуховыми устройствами, будь то слуховой аппарат или/и кохлеарный имплант, имеется реальная возможность воспринимать и обрабатывать указанные выше бинауральные признаки в стимулируемых звуковых или электрических сигналах, тем самым обеспечить необходимые условия для улучшения коммуникативных слуховых навыков
8. Укажите локализацию основных вкусовых рецепторов на языке. Укажите особенности строения и локализацию коркового конца.
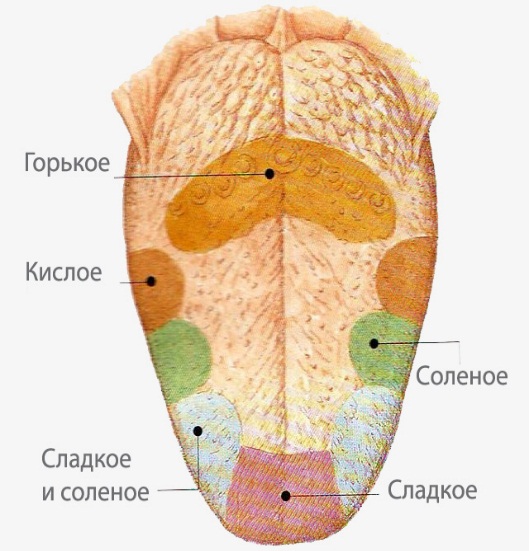
Кора — это совокупность корковых концов анализаторов. С этой точки зрения мы и рассмотрим топографию корковых отделов анализаторов, т. е. главнейшие воспринимающие участки коры полушарий большого мозга. Прежде всего рассмотрим корковые концы анализаторов, воспринимающих раздражения из внутренней среды организма.
1. Ядро двигательного анализатора, т. е. анализатора проприоцептивных (кинестетических) раздражений, исходящих от костей, суставов, скелетных мышц и их сухожилий, находится в предцентральнои извилине (поля 4 и 6) и lobulus paracentralis. Здесь замыкаются двигательные условные рефлексы. Двигательные параличи, возникающие при поражении двигательной зоны, И. П. Павлов объясняет не повреждением двигательных эфферентных нейронов, а нарушением ядра двигательного анализатора, вследствие чего кора не воспринимает кинестетические раздражения и движения становятся невозможными. Клетки ядра двигательного анализатора заложены в средних слоях коры моторной зоны. В глубоких ее слоях (V, отчасти VI) лежат гигантские пирамидные клетки, представляющие собой эфферентные нейроны, которые И. П. Павлов рассматривает как вставочные нейроны, связывающие кору мозга с подкорковыми ядрами, ядрами черепных нервов и передними рогами спинного мозга, т. е. с двигательными нейронами.В предцентральнои извилине тело человека, так же как и в задней, спроецировано вниз головой. При этом правая двигательная область связана с левой половиной тела и наоборот, ибо начинающиеся от нее пирамидные пути перекрещиваются частью в продолговатом, а частью в спинном мозге. Мышцы туловища, гортани, глотки находятся под влиянием обоих полушарий. Кроме предцентральнои извилины, проприоцептивные импульсы (мышечно-суставная чувствительность) приходят и в кору постцентральной извилины.
2. Ядро двигательного анализатора, имеющего-отношение к сочетанному повороту головы и глаз в противоположную сторону, помещается в средней лобной извилине, в премоторной области (поле 8). Такой поворот происходит и при раздражении поля 17, расположенного в затылочной доле в соседстве с ядром зрительного анализатора. Так как при сокращении мышц глаза в кору мозга (двигательный анализатор, поле 8) всегда поступают не только импульсы от рецепторов этих мышц, но и импульсы от сетчатки (зрительный анализатор, поле 17), то различные зрительные раздражения всегда сочетаются с различным положением глаз, устанавливаемым сокращением мышц глазного яблока.
Ядро двигательного анализатора, посредством которого происходит синтез целенаправленных сложных профессиональных, трудовых и спортивных движений, помещается в левой (у правшей) нижней теменной дольке, в gyrus supramarginalis (глубокие слои поля 40). Эти координированные движения, образованные по принципу временных связей и выработанные практикой индивидуальной жизни, осуществляются через связь gyrus supramarginalis с предцентральнои извилиной. При поражении поля 40 сохраняется способность к движению вообще, но появляется неспособность совершать целенаправленные движения, действовать — апраксия (праксия — действие, практика).
9. Определение боли. Понятие об эпикритической и протопатической боли.
Боль — физическое или эмоциональное страдание, мучительное или неприятное ощущение, мучение. Является одним из симптомов ряда заболеваний.
Эпикритическая боль является результатом восхождения болевой импульсации по таламокортикальному пути к нейронам соматосенсорной зоны коры большого мозга и возбуждения их. Субъективное ощущение боли формируется именно в корковых структурах.
Протопатическая боль развивается в результате активации главным образом нейронов переднего таламуса и гипоталамических структур. - Целостное ощущение боли у человека формируется при одновременном участии корковых и подкорковых структур, воспринимающих импульсацию о протопатической и эпикритической боли, а также о других видах воздействий. В коре мозга происходят отбор и интеграция информации о болевом воздействии, превращение чувства боли в страдание, формирование целенаправленного, осознанного «болевого поведения». Цель такого поведения: быстро изменить жизнедеятельность организма для устранения источника боли или уменьшения её степени, для предотвращения повреждения или снижения его выраженности и масштаба.

10. Основные элементы антиноцицептивной системы организма.
Антиноцицептивная система – это иерархическая совокупность нервных структур на разных уровнях ЦНС, с собственными нейрохимическими механизмами, способная тормозить деятельность болевой (ноцицептивной) системы.
Структура антиноцицептивной системы (АНЦ-системы)
1. АНЦ-структуры среднего, продолговатого и спинного мозга. Главные из них: серое околоводопроводное вещество (сильвиев водопровод соединяет III и IV желудочки), ядра шва и ретикулярной формации, а также желатинозная субстанция спинного мозга.
Они тормозят «ворота боли» спинного мозга, угнетают восходящий ноцицептивный поток возбуждения. Это система нисходящего тормозного контроля боли. Трансмиттерами этой тормозной системы являются опиоиды, а такжесеротонин. Более точно следует называть эти трансмиттеры модуляторами, а не медиаторами, т.к. они модулируют (изменяют) состояние нейронов-мишеней, а не передают на них возбуждение. Различия между медиаторами и модуляторами объясняются тут: Медиаторы и модуляторы.
Основные нейроны АНЦ-системы локализованы в околоводопроводном сером веществе среднего мозга. Их аксоны образуют нисходящие пути к продолговатому и спинному мозгу и восходящие пути к ретикулярной формации, таламусу, гипоталамусу, лимбической системе, базальным ганглиям и коре. Медиаторами этих нейронов являются пентапептиды: мет-энкефалин и лей-энкефалин, имеющие в качестве концевых аминокислот соответственно метионин и лейцин. Энкефалины возбуждают опиатные рецепторы. В энкефалинергических (опиатергических) синапсах опиатные рецепторы находятся на постсинаптической мембране, но эта же мембрана является пресинаптической для других синапсов - болевых, т.е. через неё должны выделяться трансмиттеры, передающие "болевое" возбуждение с одного болевого нейрона на другой.
Опиатные рецепторы являются метаботропными, они ассоциированы с аденилатциклазным биорегуляторным внутриклеточным путём и вызывают ингибирование аденилатциклазы. В результате в болевых нейронах нарушается синтез цАМФ. В итоге уменьшается вход кальция и освобождение трансмиттеров, включая медиаторы боли: субстанции P, холецистокинина, соматостатина, глутаминовой кислоты.
2. АНЦ-структуры гипоталамуса.
Они оказывают различное действие на болевую ноцицептивную систему:
1) нисходящее тормозное влияние на ноцицептивные нейроны спинного мозга;
2) восходящее тормозное влияние на таламические ноцицептивные нейроны;
3) активирующее влияние на систему нисходящего тормозного контроля (т.е. АНЦ-систему предыдущего первого уровня).
3. АНЦ-структуры второй соматосенсорной зоны коры.
Эта зона активирует АНЦ-структуры предыдущего первого и второго уровня.
Механизм работы антиноцицептивной системы
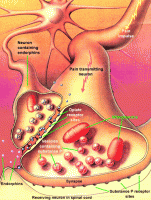
Антиноцицептивная система выделяет биологически активные эндогенные опиоидные вещества – это «внутренние наркотики». Они называются эндорфины, энкефалины, динорфины. Все они по химическому строению являются короткими пептидными цепочками, как бы кусочками белковых молекул, т. е. состоят из аминокислот. Отсюда и название: нейропептиды, опиоидные пептиды. Опиоидные — т. е. подобные по действию наркотическим веществам опийного мака.
На многих нейронах болевой системы существуют специальные молекулярные рецепторы к этим веществам. Когда опиоиды связываются с этими рецепторами, то возникает пресинаптическое и/или постсинаптическое торможение в нейронах болевой системы. Болевая ноцицептивная система тормозится и слабо реагирует на боль.
На рисунке более мелкий АНЦ-нейрон (он слева) тормозит синапс болевого нейрона и мешает ему передавать болевое возбуждение дальше.
Кроме опиоидных пептидов в регуляции боли участвуют неопиоидные пептиды, например, нейротензин. Они влияют на боль, возникающую из разных источников. Кроме того боль могут подавлять серотонин и катехоламины (норадреналин, адреналин, дофамин).
Антиноцицептивная система действует несколькими путями:
1. Срочный механизм.
Возбуждается действием болевых стимулов, использует систему нисходящего тормозного контроля. Он быстро ограничивает афферентное ноцицептивное возбуждение на уровне задних рогов спинного мозга. Этот механизм участвует в конкурентной аналгезии (обезболивании), т.е. болевая реакция подавляется, если одновременно действует другой болевой стимул.
2. Короткодействующий механизм.
Запускается гипоталамусом, вовлекает систему нисходящего тормозного контроля среднего, продолговатого и спинного мозга. Этот механизм ограничивает болевое возбуждение не только на уровне спинного мозга, но и выше, активируется стрессогенными факторами.
3. Длительнодействующий механизм.
Активируется при длительной боли. Центры его находятся в гипоталамусе. Вовлекается система нисходящего тормозного контроля. Этот механизм ограничивает восходящий поток болевого возбуждения на всех уровнях ноцицепивной системы. Этот механизм подключает эмоциональную оценку и придает эмоциональную окраску боли.
4. Тонический механизм.
Поддерживает постоянную активность антиноцицептивной системы. Центры его находятся в орбитальной и фронтальной областях коры, расположенных за лбом и глазами. Обеспечивает постоянное тормозное влияниена активность ноцицептивной структуры на всех уровнях. Важно отметить, что это происходит даже при отсутствии боли. Таким образом, с помощью антиноцицептивных структур коры больших полушарий головного мозга можно заранее подготовится и затем при действии болевого раздражителя уменьшить болезненные ощущения.
ПРАКТИЧЕСКИЕ РАБОТЫ

