 2018-01-21
2018-01-21 7141
7141Передача сигнала в химических синапсах ЦНС подобна таковой в нервно-мышечном синапсе. Однако имеется ряд отличительных особенностей, основные из которых приведены ниже.
Для возбуждения нейрона (возникновения ПД) необходимы потоки афферентных импульсов и их взаимодействие. Один пузырек (квант медиатора) содержит 110 тыс. молекул медиатора. Один ПД, пришедший в пресинаптическое окончание, обеспечивает выделение 200300 квантов медиатора. При этом возникает небольшой возбуждающий постсинаптический потенциал (ВПСП) – около 0,05 мВ (миниатюрный ВПСП). Необходимо учесть, что одновременно могут возникать не только возбуждающие, но и тормозные потенциалы. Пороговый потенциал нейрона 5–10 мВ, так как для возбуждения нейрона требуется некоторое множество импульсов. Выброс медиатора из нервного окончания обеспечивает входящий в деполяризованную терминаль Са2+, причем количество медиатора пропорционально входу ионов Са2+; четыре иона Са2+ обеспечивает выброс одного кванта медиатора. При поступлении импульсов к нейрону-мишени по различным входам в результате пространственной суммации ВПСП возникает деполяризация генераторного пункта в нейроне (аксонный холмик), которая, достигнув критической величины, обеспечивает возникновение ПД нейрона-мишени. ВПСП возникает вследствие суммарного тока в клетку и из клетки через ионные каналы различных ионов согласно их электрохимическому градиенту.
Поступивший в пресинаптическое окончание Са2+ удаляется за его пределы с помощью Са+-насоса. Прекращение действия выделившегося в синаптическую щель медиатора осуществляется частично посредством обратного захвата его пресинаптическим окончанием, частично – с помощью разрушения специальными ферментами. Например, норадреналин расщепляется моноаминоксидазой и катехолметилтрансферазой, ацетилхолин гидролизуется ацетилхолинтрансферазой, имеющейся в синаптической щели и встроенной в постсинаптическую мембрану. Прекращение действия избытков медиатора на постсинаптическую мембрану предотвращает десенситезацию – снижение чувствительности постсинаптической мембраны к действующему медиатору. Пептидные медиаторы, ферменты и другие белки, митохондрии транспортируются в пресинаптические окончания из тела клетки по аксону с помощью микротрубочек и микрофламентов, тянущихся по всей длине аксона. Из синапса ретроградно транспортируются по аксону в тело клетки вещества, регулирующие в ней синтез белка. Для этого транспорта необходимы ионы Са2+ и энергия (АТФ непрерывно ресинтезируется в аксоне).
В генерации ПД в нейронах принимают участие ионы Са2+, ток которых в клетку более медленный, чем ток Na+. В частности, в дендритах клеток Пуркинье мозжечка выявлены не только быстрые натриевые потенциалы, но и медленные кальциевые. В телах некоторых нервных клеток ПД создается преимущественно за счет Са2+.
Место возникновения генераторных ВПСП, вызывающих ПД нейрона, – тело нейрона (рис. 4.4) в непосредственной близости от места первичного возникновения ПД, располагающегося в аксонном холмике. В связи с этим некоторые авторы предлагают назвать соответствующие синапсы генераторными синапсами. Однако подавляющее большинство синапсов (в частности, в коре большого мозга – согласно расчетам, 98%) находится на дендритах и только 2% – на телах нейронов. И тем не менее площадь мембраны тела нейронов занята синапсами на 40%, дендритов – на 75%. Отношение числа синапсов к нейронам в коре большого мозга составляет 40000:1.
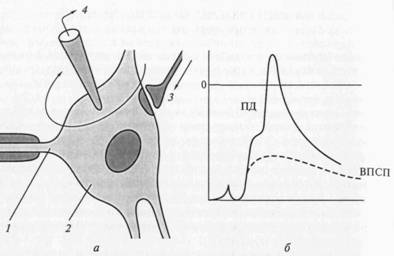
Рис. 4.4. Схема регистрации (а)и временное течение ВПСП и ПД (б) в мотонейроне спинного мозга позвоночного при одиночной стимуляции заднекорешковых волокон: 1 – начальный сегмент аксона; 2 – тело нейрона; 3 – синапс; 4 – регистрирующий электрод
Место возникновения ПД – аксонный холмик (генераторный пункт нейрона). Синапсы на нем отсутствуют, отличительной особенностью мембраны аксонного холмика является высокая ее возбудимость, в 3–4 раза превосходящая возбудимость сомадендритной мембраны нейрона, что объясняется более высокой (примерно в семь раз) концентрацией Na+-каналов на аксонном холмике. ВПСП, возникающие в любом участке сомы нейрона, за счет своего электрического поля достигают любого другого ее участка и аксонного холмика, соответственно вызывая ее деполяризацию до некоторого уровня. Это связано с тем, что постоянная длины в этой области нейрона (расстояние, на котором исходная амплитуда ВПСП уменьшается на 37%) составляет 1–2 мм, а диаметр тела нейрона равен всего 10–80 мкм. Когда величина ВПСП в результате суммации достаточна для уменьшения мембранного потенциала аксонного холмика до КУД (Екр) его мембраны, здесь генерируется ПД. Далее он распространяется, с одной стороны, антидромно на тело нейрона, с другой – ортодромно на аксон и по нему передается к другой клетке.
Роль дендритов в возникновении возбуждения. Дендритные синапсы получили название модуляторных, это связано с тем, что они удалены на значительное расстояние от генераторного пункта – аксонного холмика, поэтому их ВПСП не может вызвать достаточную деполяризацию и обеспечить возникновение ПД. Синаптический аппарат дендритов проявляет себя при одновременном поступлении возбуждения к значительному числу дендритных синапсов. При этом дендритные ВПСП изменяют потенциал мембраны сомы и аксонного холмика за счет электрического поля и модулируют возбудимость нейрона, делая ее большей или меньшей в зависимости от потока импульсов, активирующих тормозные и возбуждающие синапсы.
При возбуждении нейронов потребление О2 значительно возрастает. Источником энергии является в основном глюкоза крови, собственные небольшие запасы гликогена достаточно лишь на 3–5 мин работы нейрона.

