 2017-11-01
2017-11-01 2516
2516Розрізняють пасивний та активний транспорт. Пасивний транспорт відбувається без затрат енергії за електрохімічним градієнтом. До пасивного належать дифузія (проста та полегшена), осмос, фільтрація. Активний транспорт потребує енергії і відбувається всупереч концентраційному або електричному градієнту.
Пасивний транспорт. Дифузія може відбуватись або через ліпідний бішар, або за участю білків. Дифузія через ліпідний бішар – це проста дифузія, що залежить від концентрації речовин по обидва боки мембрани та температури. Так переходять через мембрану речовини, що розчиняються в ліпідах (О2, СО2, азот, алкоголь). Кисень проходить так легко, ніби мембрани і не існує. Через мембрану легко проходить і вода завдяки невеликому розміру її молекул.
Проста дифузія відбувається не тільки через ліпідний бішар, але й через білкові канали та “ворота” в них. Ці канали високо-селективні, транспорт через них залежить від виду іонів чи молекул, їх діаметра, форми, електричного заряду. Натрієві канали мають діаметр 0,3-0,5 нм, їх внутрішня поверхня негативно заряджена. К+канал має діаметр 0,3 нм, електричного заряду не має. Конформаційні зміни форми білкових молекул впливають на стан «воріт» (відкриті чи закриті), в Nа+-каналах вони розташовані на зовнішній, а в К+каналах – на внутрішній поверхні каналу.
Стан «воріт» контролюється двома принципово відмінними механізмами: 1) стан «воріт» залежить від різниці електричних потенціалів по обидва боки мембрани, це потенціалзалежні канали; 2) хемочутливі канали ті, білкові сполуки яких здатні зв'язуватись із деякими речовинами – лігандами. Це викликає конформаційні зміни білкової молекули і реакцію «воріт». До ліганд належить ацетилхолін.
Потенціалчутливі канали реагують за законом, що формулюється словосполученням «все або нічого». Стан «воріт» змінюється дуже швидко (мільйонні частки секунди). Зараз є точні методики дослідження потоку іонів через одиночні канали (за допомогою мікропіпеток, діаметр кінчика яких становить 1-2 мкм).
Полегшена дифузія відбувається за участю специфічного білка-переносника, що сприяє дифузії. Від простої дифузії відрізняється тим, що швидкість її збільшується тільки до певної міри, а потім залишається постійною. Таким чином транспортуються глюкоза та більшість амінокислот.
На швидкість дифузії впливають концентрація речовини по обидва боки мембрани, її проникність та ряд інших факторів. А проникність залежить від товщини мембрани, кількості каналів на одиницю площі, молекулярної маси речовини, температури.
Осмос має значення для транспорту води через мембрани, які проникні для води, але не для іонів (напівпроникні мембрани). Якщо концентрація речовини по обидва боки мембрани різна, то вона буде переходити туди, де концентрація вища. Це спостерігається доти, доки концентрація не вирівняється.
Для створення осмотичного тиску має значення не молекулярна маса речовини, а кількість молекул на одиницю об'єму розчину. Перехід води за законами осмосу відбувається дуже швидко. Визначається величина осмотичного тиску в атмосферах, у міліметрах ртутного стовпа або в міліосмолях. Ця величина у клітинній та позаклітинній рідині дорівнює 7,6 атм, або 300 мосм/кг.
Активний транспорт. Це транспорт речовин всупереч концентраційному або електричному градієнту, що відбувається із затратами енергії. Розрізняють первинний активний транспорт, що потребує енергії АТФ, та вторинний (створення за рахунок АТФ іонних концентраційних градієнтів по обидва боки мембрани, а вже енергія цих градієнтів використовується для транспорту).
Первинний активний транспорт широко використовується в організмі. Він бере участь у створенні різниці електричних потенціалів між внутрішнім та зовнішнім боками мембрани клітини. За допомогою активного транспорту створюються різні концентрації Nа+, К+, Н+, СІ- та інших іонів у середині клітини та в позаклітинній рідині.
Найкраще досліджено транспорт Nа+ та К+, Nа+,-К+-насос. Цей транспорт відбувається за участю глобулярного білка з молекулярною масою близько 100000. Білок має три ділянки для зв'язування Nа+ на внутрішній поверхні та дві ділянки для зв'язування К+ на зовнішній поверхні. Спостерігається висока активність АТФ-ази на внутрішній поверхні білка. Енергія, що утворюється при гідролізі АТФ, зумовлює конформаційні зміни білка і при цьому виводиться три іони Nа+ з клітини і вводиться у неї два іони К+. За допомогою такого насоса створюються висока концентрація Nа+ в позаклітинній рідині і висока концентрація К+ - в клітинній.
Останнім часом інтенсивно вивчаються Са+-насоси, завдяки яким концентрація Са+ у клітині в десятки тисяч разів нижча, ніж поза нею. Розрізняють Са+-насоси у клітинній мембрані і в органелах клітини (саркоплазматична сітка, мітохондрії). Са+-насоси теж функціонують за рахунок білка-переносника в мембранах. Цей білок має високу АТФ-азну активність.
Вторинний активний транспорт. Завдяки первинному активному транспорту створюється висока концентрація Nа+ за межами клітини, виникають умови для дифузії Nа+ в клітину, але разом з Nа+ інші речовини можуть увійти в неї. Цей транспорт, спрямований у один бік, називається симпортом. У іншому випадку вхід Nа+ стимулює вихід іншої речовини з клітини, це два потоки, спрямовані в різні боки,— антипорт.
Прикладом симпорту може бути транспорт глюкози або амінокислот разом з Nа+. Білок-переносник має дві ділянки для зв’язування Nа+ і для зв'язування глюкози або амінокислоти. Ідентифіковано п'ять окремих білків для зв'язування п'яти типів амінокислот. Відомі й інші види симпорту – транспорт N+ разом з СІ- у клітину, К+ і СІ- з клітини та ін.
Майже в усіх клітинах існує механізм антипорту —Nа+ переходить у клітину, а Са+ виходить з неї, або Nа+—у клітину, а Н+ - з неї.
Активно транспортуються через мембрану Мg+, Fе+, НСОз та багато інших речовин.
Піноцитоз — це один із видів активного транспорту. Він полягає в тому, що деякі макромолекули (переважно білків, макромолекули яких мають діаметр 100-200 нм) приєднуються до рецепторів мембрани. Ці рецептори специфічні для різних білків. Приєднання їх супроводжується активізацією скоротливих білків клітини — актину та міозину, які утворюють і закривають порожнину з цим позаклітинним білком і невеликою кількістю позаклітинної рідини. При цьому утворюється піноцитозний пухирець. У нього виділяються ферменти, що гідролізують цей білок. Продукти гідролізу засвоюються клітиною. Піноцитоз потребує енергії АТФ та наявності Са+ в позаклітинному середовищі.
Таким чином, є багато видів транспорту речовин через клітинні мембрани. На різних боках клітини (на апікальній, базальній, латеральній мембранах) можуть відбуватись різні види транспорту. Прикладом цього можуть бути процеси, що відбуваються в нирках при реабсорбції, а також процеси всмоктування в тонкій кишці.
 Ходжкиним виконані класичні досліди на гігантському нервовому волокні кальмара. Вирівнювали концентрацію іонів калію усередині волокна й у навколишній рідині – потенціал спокою зникав. Якщо ж волокно заповнювалося штучним сольовим розчином, близьким по складу до внутрішньоклітинної рідини, між внутрішньою і зовнішньою сторонами мембрани встановлювалася різниця потенціалів, приблизно рівна потенціалу спокою нормального волокна (50-80 мв).
Ходжкиним виконані класичні досліди на гігантському нервовому волокні кальмара. Вирівнювали концентрацію іонів калію усередині волокна й у навколишній рідині – потенціал спокою зникав. Якщо ж волокно заповнювалося штучним сольовим розчином, близьким по складу до внутрішньоклітинної рідини, між внутрішньою і зовнішньою сторонами мембрани встановлювалася різниця потенціалів, приблизно рівна потенціалу спокою нормального волокна (50-80 мв).
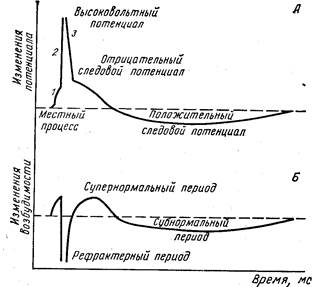
Зміни збудливості в різні фази потенціалу дії. Розвиток у нервовому чи м'язовому волокні потенціалу дії супроводжується вираженими змінами збудливості, що відбуваються по визначених фазах. Для їхнього вивчення нерв чи м'яз піддають дії двох коротких, але сильних електричних подразників, що випливають один за одним через визначені інтервали часу. Розрізняють наступні фази зміни збудливості: 1 – початкове підвищення збудливості; 2 – рефракторний період, що складається з фаз абсолютної і відносний рефрактерності; 3 – супернормальний період, чи фаза екзальтації і 4 – субнормальний період збудливості (мал. 6).
Мал. 6. Зміни збудливості нервового волокна в різні фази потенціалу дії і следовых змін мембранного потенціалу (по Моргану).
Для наочності тривалість перших двох фаз на кожної кривої трохи збільшена. Пунктирною лінією на малюнку А позначений потенціал спокою, на малюнку Б – вихідний рівень збудливості.
Початкове підвищення збудливості збігається за часом з місцевими коливаннями мембранного потенціалу і зв'язано з підвищенням проникності мембрани для всіх іонів, але особливо для іонів натрію. Виникнення піку потенціалу дії супроводжується різким зниженням збудливості тканини – станом рефрактерності. Стадія абсолютної рефрактерності збігається за часом з висхідною частиною спайка. У цей момент тканина не приходить у стан збудження навіть при дії надпорогових подразників. Виникнення абсолютної незбудливості тканини пов'язане з тим, що подальше підвищення натрієвої проникності в цей період часу неможливо. Тривалість абсолютної рефрактерності в нервовому волокні складає 0,002 з, у скелетному м’язі теплокровних – 0,005 с. Відносний рефракторний період збігається зі спадним коліном піка потенціалу дії. Збудливість у цю стадію поступово відновлюється й у відповідь на надпорогові подразники може знову виникати потенціал дії. Зазначене явище зв'язане з відбудовними процесами на мембрані і як наслідок цього можливістю підвищення натрієвої проникності на надпороговий подразник. Тривалість відносного рефрактерного періоду в нервовому волокні 0,012 з, у м'язі 0,03 с.
Відносна рефрактерність змінюється фазою екзальтації (Н. Е. Введенский), чи супернормальної збудливості. У цей період тканина відповідає виникненням збудження на подразник підпорогової (для вихідного стану) сили. Тривалість фази екзальтації в нервовому волокні 0,02 з, у м'язі 0,05 с. Період супернормальної збудливості відповідає негативному слідовому потенціалу.
Слідом за фазою екзальтації виникає стадія субнормальної збудливості, яка співпадає з позитивним слідовим потенціалом. Збудливість у цю фазу незначно знижена в порівнянні з вихідним її рівнем.

