 2017-11-01
2017-11-01 776
776А і В – електроди осцилографа; О – осцилограф. Стрілкою показані місце і момент нанесення подразнення; + і – характеризують електричний заряд нерва в даний момент. Зміна заряду приводить до зміни положення променя на екрані осцилографа.
 Мал. 4. Схема потенціалу дії, зареєстрованого за допомогою внутрішньоклітинного мікроелектрода. а – місцеві коливання мембранного потенціалу; d – висхідна частина піку потенціалу дії (фаза деполяризації); c – спадна частина піка потенціалу дії (фаза реполяризації); d – негативний слідовий потенціал; e – позитивний слідовий потенціал. Стрілкою показаний момент нанесення подразнення.
Мал. 4. Схема потенціалу дії, зареєстрованого за допомогою внутрішньоклітинного мікроелектрода. а – місцеві коливання мембранного потенціалу; d – висхідна частина піку потенціалу дії (фаза деполяризації); c – спадна частина піка потенціалу дії (фаза реполяризації); d – негативний слідовий потенціал; e – позитивний слідовий потенціал. Стрілкою показаний момент нанесення подразнення.
Реєстрація потенціалу дії за допомогою електронних підсилювачів і мікроелектродів дозволила з'ясувати досить складну його структуру (мал. 4).
У потенціалі дії розрізняють місцеві коливання мембранного потенціалупік потенціалу дії і слідові потенціали – негативний і позитивний. Пік потенціалу дії представлений висхідною (В) і спадною (С) гілками. Під час висхідної частини піку потенціалу дії заряд мембрани, швидко знижується до нуля (деполяризація) і виникає різниця потенціалів протилежного знака. У зв'язку з цим висхідна частина піку одержала назву фази деполяризації. У момент спадної частини піку відбувається відновлення вихідного рівня поляризації (електричного стану) мембрани. Унаслідок цього цю частину потенціалу дії називають фазою реполяризації. Слідові потенціали зв'язані з відбудовними процесами, що повільно розвиваються в нервових і м'язових волокнах по закінченні процесу збудження. Тривалість потенціалу дії в нервових і м'язових волокнах складає. 0,1-5 мс, при цьому фаза реполяризації завжди триваліше фази деполяризації. Слідові потенціали мають низьку амплітуду – кілька мілівольтів і більш тривалий, чим потенціал дії. Тривалість слідових потенціалів може коливатися в дуже широких межах – від декількох мілісекунд до декількох десятків і навіть сотень мілісекунд.
Походження потенціалу спокою і потенціалу дії. Запропоновано багато теорій, що пояснюють походження біопотенціалів. Найбільше повно експериментально обґрунтована мембранна теорія, запропонована німецьким дослідником Бернштейном (1902, 1912). У сучасний період ця теорія модифікована й експериментально розроблена Ходжкиним, Хаксли, Катцем (1949-1952).
Встановлено, що в основі біоелектричних явищ лежить нерівномірний розподіл (асиметрія) іонів у цитоплазмі клітини і навколишньому середовищі. Так, протоплазма нервових і м'язових клітин містить у 30-50 разів більше іонів калію, у 8-10 разів менше іонів натрію й у 50 разів менше іонів хлору, чим позаклітинна рідина. Крім того, до складу цитоплазми клітини входять органічні аніони (крупномолекулярні з'єднання, що несуть негативний заряд), що відсутні в позаклітинному середовищі.
Прихильники мембранної теорії основною причиною іонної асиметрії вважають наявність клітинної мембрани зі специфічними властивостями.
Будова і функції клітинної мембрани (плазмолеми). Плазмолема складається з двох шарів молекул ліпідів (переважно фосфоліпідів) – ліпідного бішару та білків, які пронизують бішар ліпідів наскрізь – інтегральні білки – або виступають з нього назовні чи в середину клітини – периферичні білки. Більшість із цих білків, що ніби плавають у ліпідах,—глікопротеїни. На поверхні білків, що виступають назовні, можуть бути фіксовані молекули вуглеводів (мал. 5).
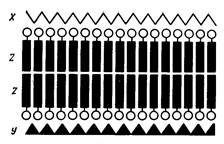 Мал. 5. Схема молекулярної структури клітинної мембрани. Показано бімолекулярний ліпідний шар – Z. (кружками позначені полярні групи фосфоліпідів) і два неліпідних монослоя: зовнішній – мукополісахаридний, X – внутрішній – білковий, Y.
Мал. 5. Схема молекулярної структури клітинної мембрани. Показано бімолекулярний ліпідний шар – Z. (кружками позначені полярні групи фосфоліпідів) і два неліпідних монослоя: зовнішній – мукополісахаридний, X – внутрішній – білковий, Y.
Плазмолема дуже тонка (завтовшки 7,5-10 нм) і еластична. Основні її функції: створення бар'єру між середовищем клітини та навколишньою міжклітинною рідиною; участь у підтриманні внутрішньої структури клітини, її цитоскелета; рецепція різних медіаторів, гормонів та інших фізіологічно активних речовин; зміна проникності мембрани для різних речовин; створення електричних потенціалів по обидва боки мембрани. Важливі також антигенні властивості цих мембран.
Завдяки бар'єрній функції створюється різниця у складі рідини як у власне клітині, так і за її межами.
У стані відносного фізіологічного спокою мембрана має підвищену проникність для іонів калію, проникність же її для іонів натрію різко знижена.
Таким чином, особливості проникності клітинної мембрани, а також розміри самих іонів є однієї з причин, що забезпечують асиметрію розподілу іонів по обох сторонах клітинної мембрани. Іонна жасиметрія – одна з основних причин виникнення потенціалу спокою, при цьому ведуча роль належить нерівномірному розподілу іонів калію.
Механізм виникнення потенціалу дії значно складніше. Основна роль у виникненні струмів дії належить іонам натрію. При дії подразника граничної сили проникність мембрани клітини для іонів натрію зростає в 500 разів і перевищує проникність для іонів калію в 10-20 разів. У зв'язку з цим натрій лавиноподібно спрямовується в клітку, що приводить до перезарядження клітинної мембрани. Зовнішня поверхня заряджається негативно стосовно внутрішнього. Відбувається деполяризація клітинної мембрани, що супроводжується реверсією мембранного потенціалу. Під реверсією мембранного потенціалу розуміють та кількість мілівольтів (мв), на яке потенціал дії перевищує потенціал спокою. Відновлення вихідного рівня мембранного потенціалу (реполяризація) здійснюється за рахунок різкого зниження натрієвої проникності (інактивація) неактивного переносу іонів натрію з цитоплазми клітини в навколишнє середовище.
Доказу натрієвої гіпотези потенціалу дії також були отримані Ходжкиним. Дійсно, якщо потенціал дії має натрієву природу, то, варіюючи концентрацію іонів натрію, можна змінити величину потенціалу дії. Виявилося, що при заміні 1/3 морської води, що є нормальним навколишнім середовищем для гігантського аксона кальмара, на ізотонічний розчин декстрози, тобто при зміні концентрації натрію в навколишнім середовищі на 2/3, потенціал дії зменшується наполовину.
Таким чином, виникнення біопотенціалів є функцією біологічної мембрани, що володіє виборчою проникністю. Величина потенціалу спокою і потенціалу дії обумовлюється іонною асиметрією в системі клітина-середовище.
Існує градієнт (міра зростання або спадання в просторі якоїсь фізичної величини на одиницю довжини) концентрації іонів та молекул по обидва боки плазмолеми. Цей градієнт пояснюється тим, що транспортуються іони та молекули через мембрану по-різному.








