 2017-11-01
2017-11-01 769
769Мобільні елементи LINE довжиною приблизно 6 тис. пар основ містять принаймні два гени – зворотної транскриптази та ендонуклеази і невеликі специфічні послідовності на кінцях. Процес переміщення копії елемента починається так само, як і у випадку ретропозонів із транскрипції РНК-полімеразою ІІ (рис. 17). Після трансляції в цитоплазмі синтезовані білки зв’язуються з мРНК, цей комплекс повертається до ядра, де й відбувається зворотна транскрипція: ендонуклеаза здійснює одноланцюговий розріз у геномній ДНК, 3′-кінець у місці розрізу використовується як праймер. Після синтезу другого ланцюга ДНК нова копія елемента остаточно вбудовується в геном.
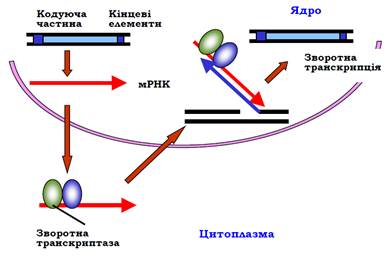
Рис. 17. Механізм переміщення мобільного елемента LINE.
Синтез мРНК при транскрипції елемента LINE, як і для більшості інших еукаріотичних мРНК, закінчується на polyA-сигналі. Цей polyA-сигнал є слабким, що дозволяє елементу вбудовуватись в інтрони звичайних генів без особливих перешкод для експресії цих генів: система процесингу часто не помічає слабкий polyA-сигнал. Елементи LINE є, відповідно, найпоширенішими мобільними елементами в геномі вищих еукаріотів (~20 % геному людини представлено послідовностями LINE) і основним джерелом зворотної транскриптази у клітинах. З часом накопичуються мутації й окремі елементи LINE стають неактивними (тобто вже не кодують зворотну транскриптазу), але переміщуються, використовуючи білки, що кодуються активними елементами. Найчастіше в геномі людини зустрічається елемент типу LINE, котрий позначається як L1 (кілька сотень тисяч копій).
Іноді мобільні елементи є не просто ділянками «егоїстичної ДНК», що автономно розмножуються в геномі, а виконують певні конкретні біологічні функції. Наприклад, у дрозофіли відсутня теломеразна система й елемент LINE певного типу виконує функцію подовження кінців хромосом після реплікації: зворотна транскриптаза виступає як теломераза, мРНК мобільного елемента – як теломеразна матричнаРНК. Цікаво, що послідовності ДНК гена теломерази та елементів LINE характеризуються високою гомологією: цілком можливо, що теломеразна система походить від мобільних елементів LINE.
Мобільні елементи SINE – короткі (100–300 пар основ) елементи послідовності, які не кодують білків і містять лише промотор для РНК-полімерази ІІІ. Процес їхнього переміщення є таким самим, як зображено на рис. 17: елементи SINE «паразитують» на системі LINE, використовуючи її ферменти. На відміну від усіх інших мобільних елементів, елементи SINE сконцентровані в ділянках геному, збагачених на гени. Імовірною причиною цієї закономірності є те, що РНК продукт SINE має спорідненість до певної протеїнкінази, яка гальмує процес трансляції. Відповідно, зв’язування РНК із протеїнкіназою блокує останню, що приводить до загального підвищення рівня білкового синтезу. Таким чином, підвищення рівня транскрипції SINE разом з активацією інших генів стимулює трансляцію.
КОНТРОЛЬНІ ЗАПИТАННЯ
1. Як саме починається гомологічна рекомбінація? Як здійснюється точна репарація дволанцюгових розривів за рекомбінаційним механізмом?
2. Що таке гетеродуплекс? Яку роль відіграє білок recA при гомологічній рекомбінації?
3. Дайте визначення структури Холідея. В якій формі вона реалізується під час гомологічної рекомбінації? За рахунок чого стабілізується така форма?
4. За яким механізмом відбувається міграція гілки під час гомологічної рекомбінації?
5. Як здійснюється поділ структури Холідея під час гомологічної рекомбінації? З якою імовірністю відбувається обмін ділянками між двома гомологічними молекулами ДНК і від чого залежить реалізація такого обміну?
6. Що відбувається з гетеродуплексом після завершення гомологічної рекомбінації?
7. Чим відрізняється сайтс-пецифічна рекомбінація від гомологічної?
8. Що таке незаконна рекомбінація?
9. Як побудовані кластери імуноглобулінових генів? Які події при дозріванні імуноглобулінового гена можна віднести до сайт-специфічної рекомбінації?
10. Якими механізмами забезпечується необмежене зростання розмаїття послідовностей імуноглобулінових генів при їхньому дозріванні?
11. Яка різниця між реплікативним і нереплікативним механізмами переміщення ДНК-транспозонів?
12. Яка різниця між механізмами переміщення LTR-ретропозонів і мобільних елементів типу LINE?
13. Чим різняться мобільні елементи типів LINE і SINE?
РЕКОМЕНДОВАНА ЛІТЕРАТУРА
1. Boeke, J.D., Chapman, K.B. Retrotransposition mechanisms // Curr.Opin. Cell Biol. – 1991. – Vol. 3. – P. 502 – 507.
2. Grindley, N.G.F., Leschziner, A.E. DNA transposition: from a black boxto a color monitor // Cell. – 1995. – Vol. 83. – P. 1063 – 1066.
3. Haniford, D.B., Chaconas G. Mechanistic aspects of DNA transposition// Curr. Opin. Genet. Dev. – 1992. – Vol. 2. – P. 698 – 704.
4. International Human Genome Sequencing Consortium. Initial sequencingand analysis of the human genome // Nature. – 2001. – Vol. 409. –– P. 860 – 921.
5. Kazazian, H.H., Moran, J.V. The impact of L1 retrotransposons on thehuman genome // Nature Gen. – 1998. – Vol. 19. – P. 19 – 24.
6. Kuzimov, A. Unraveling the late stages of recombinational repair: metabolismof DNA junctions in E. coli // BioEssays. – 1996. – Vol. 18.– P. 757 – 765.
7. Labhart, P. Nonhomologous DNA end joining in cellfree systems// Eur. J. Biochem. – 1999. – Vol. 265. – P. – 849 – 861.
8. Landy, A. Dynamic, structural, and regulatory aspects of lambda sitespecificrecombinations // Annu. Rev. Biochem. – 1989. – Vol. 58. – P. 913 – 949.
9. Levin, K.L. It’s prime time for reverse transcriptase // Cell. – 1997. – Vol. 88. – P. 5 –8.
10. Sadofsky, M.J. The RAG proteins in V(D)J recombination: more thanjust a nuclease // Nucl. Acids Res. – 2001. – Vol. 29. – P. 1399 – 1409.
11. Singer, M. F. LINE1: a human transposable element // Gene. – 1993. – Vol. 135. – P. 183 – 188.
12. Stahl, F. Meiotic recombination in yeast: coronation of the doublestrandbreak repair model //Cell. – 1996. – Vol. 87. – P. 965 – 968.
13. West, S.F. Enzymes and molecular mechanisms of genetic recombinations// Annu. Rev. Biochem. – 1992. – Vol. 61. – P. 603 – 640.
14. West S. Processing of recombination intermediates by the Ruv ABCproteins // Annu. Rev. Genet. – 1997. – Vol. 31. – P. 213 – 244.








