 2014-01-31
2014-01-31 1697
1697Конечный продукт цикла Кальвина фруктозо-6-фосфат путём изомеризации очень легко превращается в глюкозо-6-фосфат под действием фермента глюкозофосфатизомеразы:
 |
фруктозо-6-фосфат глюкозо-6-фосфат
Вследствие того, что реакции изомеризации сопровождаются небольшим изменением свободной энергии, они в большинстве своём обратимы.
Под действием фермента маннозофосфатизомеразы фруктозо-6-фосфат изомеризуется в маннозо-6-фосфат:
 |
фрукто-6-фосфат маннозо-6-фосфат
 |
В результате действия соответствующих мутаз, осуществляющих внутримолекулярный перенос фосфатной группы, из глюкозо-6-фосфата образуется глюкозо-1-фосфат, а из маннозо-6-фосфата – маннозо-1-фосфат:
глюкозо-6-фосфат глюкозо-1-фосфат
 |
маннозо-6-фосфат маннозо-1-фосфат
В нефотосинтезирующих клетках растений и у гетеротрофных организмов фруктозо-6-фосфат и глюкозо-6-фосфат включаются в реакции гликолиза или пентозофосфатного цикла. В ходе реакций гликолиза фосфорилированная гексоза расщепляется на две триозы – 3-фосфогли-цериновый альдегид и фосфодиоксиацетон, из которых при обращении реакций гликолиза может снова синтезироваться гексоза. В пентозофосфатном цикле образуется примерно такой же набор фосфорнокислых производных моносахаридов, как и в цикле Кальвина. Таким образом, мы видим, что в гликолитических реакциях осуществляются взаимные превращения гексоз и триоз на уровне их фосфорнокислых эфиров, а в пентозофосфатном цикле и цикле Кальвина происходят взаимные превращения фосфорилированных форм триоз, пентоз, эритрозы, глюкозы и фруктозы.
Включение свободных моносахаридов в различные реакции их превращений осуществляется путем фосфорилирования. В качестве донора фосфатных групп для реакций фосфорилирования чаще всего используется АТФ. Ферменты, катализирующие перенос фосфатных групп от АТФ на моносахариды, как мы уже знаем, называют киназами.
Фосфорилирование глюкозы и маннозы катализирует фермент гексокиназа с образованием глюкозо-6-фосфата и маннозо-6-фосфата.
По аналогичному механизму фруктокиназа катализирует образование фруктозо-1-фосфата из фруктозы, а фермент галактокиназа синтезирует галактозо-1-фосфат из галактозы.
Образование свободных моносахаридов из их фосфорилированных форм катализируют ферменты гидролитического типа – фосфатазы, которые отщепляют от фосфатов моносахаридов остатки фосфорной кислоты. Гидролиз глюкозо-6-фосфата происходит с участием глюкозо-6-фосфатазы, галактозо-1-фосфата – под действием галактозо-1-фосфатазы, фруктозо-6-фосфата – фруктозо-6-фосфатазы и т.д.
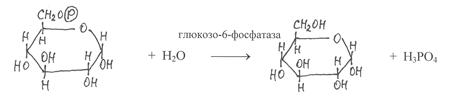 |
глюкозо-6-фосфат глюкоза
Значительно сложнее происходят взаимопревращения галактозы и других моносахаридов. В этих реакциях в качестве промежуточных продуктов образуются нуклеозиддифосфатпроизводные соответсвующих моносаха-ридов. На первом этапе таких превращений галактоза фосфорилируется с участием фермента галактокиназы, в результате образуется галактозо-1-фосфат:
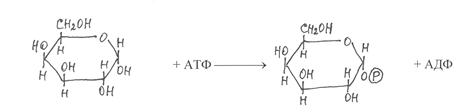 |
галактоза галактозо-1-фосфат
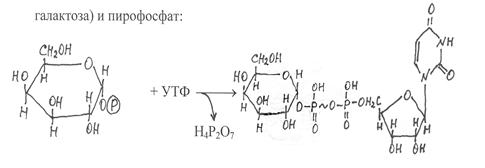 |
На следующей стадии галактозо-1-фосфат взаимодействует с уридинтрифосфатом (УТФ). Эту реакцию катализирует фермент галактозо-1-фосфатуридилилтрансфераза, под действием которого обра-
галактозо-1-фосфат УДФ-галактоза
зуется нуклеотидное производное галактозы – уридиндифосфат-галактоза (УДФ-галактоза) и пирофосфат.
В дальнейшем УДФ-галактоза изомеризуется в УДФ-глюкозу под действием специфической НАД-зависимой 4-эпимеразы:
 |
УДФ-галактоза УДФ-глюкоза
После гидролитического расщепления УДФ-глюкоза распадается на два продукта – глюкозо-1-фосфат и уридинмонофосфат (УМФ):
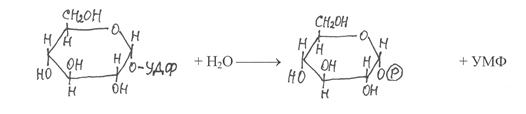 |
УДФ-глюкоза глюкозо-1-фосфат
Глюкозо-1-фосфат может далее изомеризоваться в глюкозо-6-фосфат, а глюкозо-6-фосфат – во фруктозо-6-фосфат. Таким образом, посредством указанных реакций галактоза может превращаться во фруктозо-6-фосфат, который включается в реакции дыхания, или в глюкозо-6-фосфат, способный превращаться в продукты пентозофосфатного цикла.
Возможен также синтез галактозо-1-фосфата из глюкозо-1-фосфата, так как в клетках организмов содержится фермент глюкозо-1-фосфатури-дилилтрансфераза, катализирующий образование УДФ-глюкозы из глюкозо-1-фосфата и УТФ:
глюкозо-1-фосфат + УТФ ¾¾® УДФ-глюкоза + Н4Р2О7
Затем УДФ-глюкоза под действием 4-эпимеразы изомеризуется в УДФ-галактозу, при гидролизе которой образуется галактозо-1-фосфат:
УДФ-галактоза + Н2О ¾¾® галактозо-1-фосфат + УМФ
Взаимопревращения гексоз и пентоз осуществляются в пентозофосфатном цикле и цикле Кальвина. Важное значение для этих реакций имеют ферменты транскетолаза и трансальдолаза, а в пентозо-фосфатном цикле – ещё и фермент фосфоглюконатдегидрогеназа, ката-лизирующий окислительное декарбоксилирование 6-фосфоглюконовой кислоты с образованием рибулозо-5-фосфата. Этот фермент фактически осуществляет превращение гексозы в пентозу. Во взаимных превращениях пентоз также участвуют ферменты рибулозо-фосфатэпимераза и рибозофосфатизомераза, поддерживающие динамическое равновесие между рибулозо-5-фосфатом, с одной стороны, и ксилулозо-5-фосфатом и рибозо-5-фосфатом, с другой стороны.
Ксилоза и арабиноза синтезируются также из гексоз, но другим путём. При этом в качестве промежуточных продуктов образуются нуклеотидные производные глюкуроновой и галактуроновой кислот. На первом этапе осуществляется ситез УДФ-глюкозы из глюкозо-1-фосфата и
УТФ, а затем под действием фермента УДФ-глюкозодегидрогеназы (1.1.1.22) УДФ-глюкоза окисляется в УДФ-глюкуроновую кислоту:
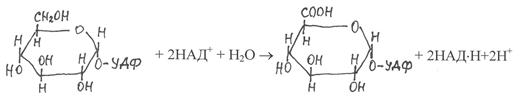 |
УДФ-глюкоза УДФ-глюкуроновая
кислота
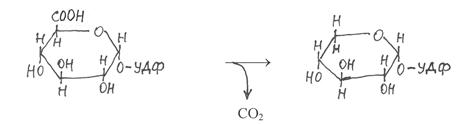 |
Затем УДФ-глюкуроновая кислота подвергается декарбоксили-рованию и превращению в пиранозную форму УДФ-ксилозы:
УДФ-глюкуроновая УДФ-ксилоза
кислота
Полученная таким путём УДФ-ксилоза используется в процессе синтеза ксиланов.
 |
По аналогичному механизму осуществляется синтез УДФ-арабинозы из УДФ-галактозы, при этом в качестве промежуточного продукта образуется УДФ-галактуроновая кислота. УДФ-арабиноза так же, как и УДФ-ксилоза, не накапливается в растительных тканях, а используется для синтеза арабанов. Кроме того, возможны взаимные превращения УДФ-глюкуроновой и УДФ-галактуроновой кислот, а также пираназных форм УДФ-ксилозы и УДФ-арабинозы под действием соответствующих 4-эпимераз.
УДФ-глюкуроновая кислота УДФ-галактуроновая кислота
УДФ-галактуроновая кислота является основным источником галактуроновой кислоты для синтеза пектиновых веществ, а УДФ-глюкуроновая кислота участвует в синтезе ксиланов (в качестве ответвлений), полиуренидов, аскорбиновой кислоты.
Превращение моносахаридов в спирты. У растений, грибов и водорослей важную роль в углеводном обмене играют спирты – сорбит, маннит, дульцит.
Дульцита много содержится в листьях ряда растений, особенно в листьях бересклета. Сорбита много накапливается в плодах и ягодах. Очень много маннита в грибах и водорослях, а также в некоторых растительных продуктах. Синтез этих спиртов осуществляется в результате восстановления соответствующих моносахаридов. Наиболее хорошо изучена реакция образования маннита из фруктозы. Эту реакцию катализирует фермент маннитолдегидрогеназа (1.1.1.138):
|
Спирты, синтезируемые в результате восстановления моносахаридов, выполняют функцию резервных углеводов. Они очень легко превращаются в соответствующие моносахариды.
8.3. Синтез и распад олигосахаридов и полисахаридов.
Наиболее распространенный олигосахарид растений – сахароза, который синтезируется только в клетках растений и выполняет в них роль транспортной формы, а также может накапливаться в качестве запасного вещества в корнеплодах сахарной свеклы, сахарном тростнике, овощах, плодах и ягодах. В листьях растений синтез сахарозы происходит в цитоплазме фотосинтезирующих клеток из УДФ-глюкозы и фруктозо-6-фосфата, образующегося в реакциях цикла Кальвина.
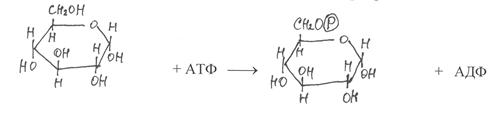 |
На первом этапе синтеза сахарозы a-глюкоза подвергается активированию путём фосфорилирования от АТФ под действием фермента гексокиназы. В результате реакции образуется глюкозо-6-фосфат и АДФ:
a-глюкоза глюкоза-6-фосфат
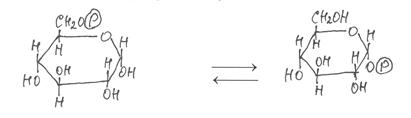 |
Затем глюкозо-6-фосфат изомеризеутся в глюкозо-1-фосфат с участием фермента фосфоглюкомутазы:
глюкозо-6-фосфат глюкозо-1-фосфат
В следующей реакции глюкозо-1-фосфат взаимодействует с уридинтрифосфатом (УТФ), при этом образуются уридиндифосфатглюкоза (УДФ-глюкоза) и пирофосфорная кислота. Реакцию катализирует фермент глюкозо-1-фосфатуридилилтрансфераза (2.7.7.9):
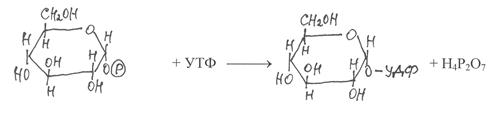 |
глюкозо-1-фосфат УДФ-глюкоза
После этого из УДФ-глюкозы с участием фруктозо-6-фосфата осуществляется синтез сахарозофосфата под действием фермента сахарозофосфат-УДФ-глюкозилтрансферазы (2.4.1.14):
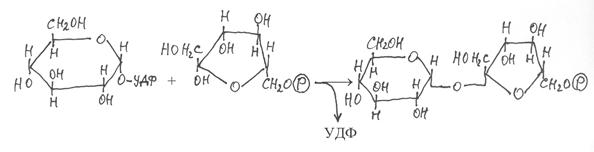 |
УДФ-глюкоза фруктозо-6-фосфат сахарозофосфат
С участием фермента сахарозофосфатазы сахарозофосфат гидролизуется с образованием сахарозы и фосфорной кислоты:
сахарозофосфат + Н2О ¾® сахароза + Н3РО4
Таким образом, для синтеза сахарозы затрачивается энергия макро-эргических связей АТФ и УТФ, необходимых для активирования a-глю-козы, а также энергия биоэнерггетических продуктов световой фазы фотосинтеза, которая потребляется в реакциях цикла Кальвина при образовании фруктозо-6-фосфата.
В нефотосинтезирующих клетках растений (корнеплодов, клубней картофеля и земляной груши, зародышей пшеницы и кукурузы, семян гороха и др.) найден фермент сахарозо-УДФ-глюкозилтрансфераза (2.4.1.13), катализирующий синтез сахарозы из УДФ-глюкозы и фруктозы в соответсвии со следующей реакцией:
УДФ-глюкоза + фруктоза D сахароза + УДФ
Следует отметить, что при высокой концентрации УДФ данный фермент может катализировать и обратную реакцию образования УДФ-глюкозы и фруктозы из сахарозы. С помощью такой реакции, например, происходит включение транспортной формы углеводов–сахарозы в биохимические превращения, имеющие место в акцепторных клетках растений.
Другой путь включения сахарозы в обмен веществ организма – её гидролиз под действием фермента b-фруктофуранозидазы, который даёт свободные формы моносахаридов глюкозы и фруктозы:
сахароза + Н2О ¾® глюкоза + фруктоза
Фермент b-фруктофуранозидаза (или инвертаза) содержится в клетках растений, животных, грибов. В клетках бактерий найден также фермент сахарозофосфорилаза, который способен превращать сахарозу во фруктозу и глюкозо-1-фосфат:
сахароза + Н3РО4 ¾®глюкозо-1-фосфат + фруктоза
Образующийся в этой реакции глюкозо-1-фосфат может затем непосредственно включиться в реакции анаэробного дыхания.
Синтез полисахаридов катализируют ферменты гликозилтрансфе-разы, которые осуществляют перенос остатков соответствующих моносахаридов, связанных с нуклеозиддифосфатными группировками, на акцептор, представляющий собой олигосахарид, который включает 2-4 соединённых О-гликозидными связями моносахаридных остатка. При этом могут синтезироваться полимеры, имеющие линейную (цепочечную) структуру или разветвлённые молекулы, состоящие как из одинаковых, так и из разных моносахаридных остатков. Многие гликозилтрансферазы представлены белками, которые связаны в определённых участках с внутриклеточными мембранами.
Синтез крахмала. Крахмал в растительных тканях представлен двумя полисахаридами амилозой и амилопектином. Синтез амилозы происходит в 3 этапа. Вначале осуществляется активирование a-глюкозы путём фосфорилирования и образования аденозиндифосфатглюкозы (АДФ-глюкозы) под действием фермента АДФГ-пирофосфорилазы:
гексокиназа
глюкоза + АТФ ¾¾¾® глюкозо-6-фосфат + АДФ
фосфоглюко-
глюкозо-6-фосфат ¾¾¾® глюкозо-1-фосфат
мутаза
АДФГ-пиро-
глюкозо-1-фосфат + АТФ ¾¾¾® АДФ-глюкоза + Н4Р2О7
фосфорилаза
На следующем этапе с участием АДФ-глюкозы под действием фермента глюкозилтрансферазы синтезируется олигосахарид, состоящий из 2-4 остатков глюкозы, соединённых a(1®4)-связями. Фермент глюкозилтрансферазу очень часто называют D-ферментом. Образующийся под действием D-фермента олигосахарид далее служит акцептором для присоединения глюкозных остатков от АДФ-глюкозы при синтезе полимера.
Образование цепочечных структур молекул амилозы катализирует фермент АДФГ-крахмалглюкозилтрансфераза (2.4.1.21). Реакция протека-ет по следующей схеме:
(глюкоза)n + АДФ-глюкоза ¾® (глюкоза)n+1 + АДФ
первичный акцептор промежуточный продукт
полимеризации
В этой реакции с помощью фермента остаток глюкозы от АДФ-глюкозы переносится на первичный акцептор, в результате чего его глюкозная цепь удлиняется на один остаток. Затем полученный продукт становится акцептором следующего остатка глюкозы и так продолжается присоединение глюкозных остатков от АДФ-глюкозы на соответствующий промежуточный акцептор, пока не закончится полный синтез молекулы амилозы.
В ходе синтеза амилозы образуется длинная цепь до 300 глюкозных остатков, соединённых О-гликозидными a(1®4)-связями. При этом следует отметить, что остатки глюкозы в процессе синтеза крахмала всегда присоединяются к нередуцирующим концам полисахаридной цепи акцептора (т.е. со стороны НО-группы четвёртого углеродного атома глюкозы).
Синтез a(1®6)-связяей в молекулах амилопектина, за счёт которых образуются разветвлённые молекулы, осуществляется с участием так называемого Q-фермента, который по современной номенклатуре ферментов получил название a-глюкантрансферазы (2.4.1.18). Q-фермент способен катализировать перенос определённого участка полиглюкозной цепи на НО-группу шестого углеродного атома одного из глюкозных остатков прилегающей и параллельно расположенной полисахаридной цепи. Расстояние между ответвлениями в цепи зависит от природы фермента.
Донорорм глюкозных остатков для синтеза крахмала может также служить УДФ-глюкоза, но при этом скорость реакции очень сильно замедляется. Однако в клетках животных организмов основным источником глюкозных остатков для построения молекул гликогена (аналога крахмала) служит УДФ-глюкоза.
Распад крахмала. Распад молекул крахмала может происходить путём гидролиза или фосфоролитических реакций. Гидролитическое расщепление a(1®4)-связей в молекулах крахмала катализируют амилазы: a-амилаза (3.2.1.1), b-амилаза (3.2.1.2), глюкоамилаза (3.2.1.3).
a-Амилазы действуют на a(1®4)-связи между точками ветвления и способны расщеплять молекулы амилопектина на более мелкие фрагменты, представляющие собой низкомолекулярные полисахариды – декстрины. Для проявления каталитической активности a-амилаз необходимо присутствие в реакционной среде хлорид-ионов, которые служат активаторами фермента. Без участия a-амилаз невозможно полное гидролитическое расщепление молекул амилопектина.
Под действием b-амилаз происходит гидролитическое расщепление a(1®4)-связей на концах полисахаридных цепей целых молекул или декстринов с образованием b-мальтозы. Действие этих ферментов прекращается при достижении точек ветвления молекул крахмала, в которых глюкозные остатки соединены a(1®6)-связями.
Глюкоамилазы так же, как и b-амилазы, катализируют гидролиз a(1® 4)-связей на концах полисахаридных цепей, но в результате действия этих ферментов образуются молекулы глюкозы.
Гидролитическое расщепление a(1®6)-связей в точках ветвления молекул амилопектина катализируют R-ферменты, которые называют амилопектин-1,6-глюкозидазами (3.2.1.9)
Под действием всего набора амилолитических ферментов крахмал гидролизуется с образованием мальтозы и глюкозы. Однако на мальтозу также действуют ферменты, относящиеся к гидролазам, a-глюкозидазы (3.2.1.20), которые расщепляют молекулы мальтозы с образованием глюкозы. Схематически действие гидролитических ферментов на молекулу крахмала показано на рисунке 38.
Препараты, содержащие амилолитические ферменты, используются в производстве хлеба, пива, пищевого спирта, а также в качестве кормовых добавок в животноводстве для улучшения переваривания крахмала, содержащегося в кормах.
Фосфоролитическое расщепление молекул крахмала катализируют ферменты a-глюканфосфорилазы (2.4.1.1). Под действием этих ферментов осуществляется перенос глюкозных остатков от молекул крахмала на фосфорную кислоту, при этом в качестве основного продукта реакции образуется глюкозо-1-фосфат, который далее может быть использован для синтеза УДФ-глюкозы или включаться в анаэробную стадию дыхания. Реакции фосфоролиза крахмала проходят по следующей схеме:
(глюкоза)n + Н3РО4 ¾® глюкозо-1-фосфат + (глюкоза)n-1
крахмал

