 2013-12-28
2013-12-28 963
963Глава 8. Структура вымираний морских биот в меловом периоде
Катастрофы и история Земли: Новый униформизм: Пер. С англ./Под ред. У. Берггрена и Дж. Ван Кауверинга. - М.: Мир, 1986.- 471 с.
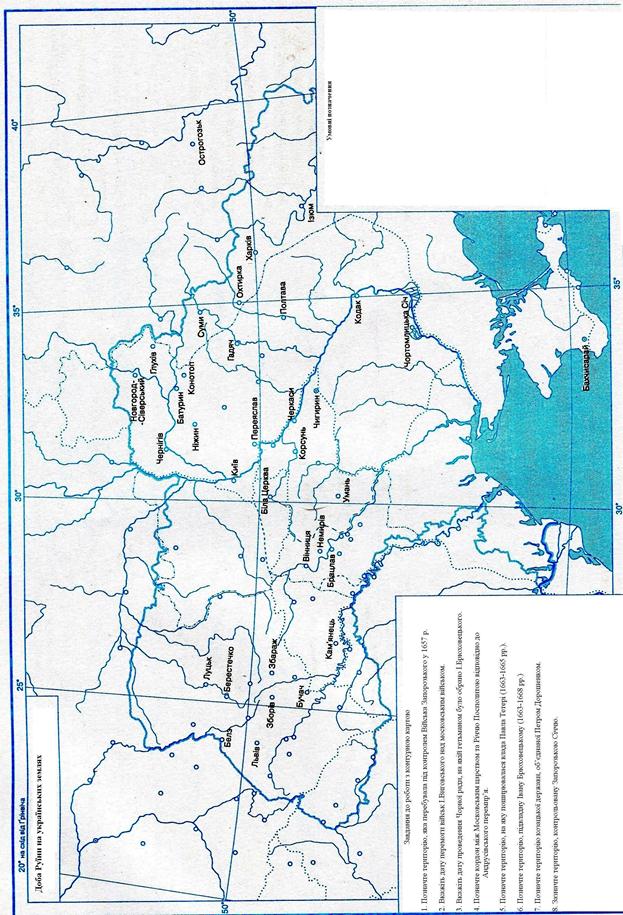
Доба Руїни на українських землях
Контурна карта № 4.
Завдання до роботи з контурною картою
- Позначте територію, яка перебувала під контролем Війська Запорозького у 1657 р.
- Вкажіть дату перемоги військ І.Виговського над московським військом.
- Вкажіть дату проведення Чорної ради, на якій гетьманом було обрано І.Брюховецького.
- Позначте кордон між Московським царством та Річчю Посполитою відповідно до Андрусівського перемир’я.
- Позначте територію, на яку поширювалася влада Павла Тетері (1663-1665 рр.).
- Позначте територію, підвладну Івану Брюховецькому (1663-1668 рр.)
- Позначте територію козацької держави, об’єднаної Петром Дорошенком.
- Зазначте територію, контрольовану Запорозькою Січчю.
Умовні позначення
| територія під контролем Війська Запорозького у 1657 р. | |
| перемога І.Виговського над московським військом |
| ● | місце Чорної ради |
| ║ | російсько-польський кордон за Андрусівським перемир’ям |
| територія, підвладна Павлу Тетері | |
| територія, підвладна Івану Брюховецькому | |
| козацька держава Петра Дорошенка |
| територія, підконтрольна Запорозькій Січі |
Допоміжні матеріали
- Історія України. Ілюстрований атлас. – К.: Інститут передових технологій, 2008. – С. 24, 25
- ХДУ. Історія України. https://vk.com/club47737829
Фотоальбом "Козацька доба" – історичні карти "Руїна 1657-1667 рр." та "Руїна 1667-1687 рр.".
Э. Дж. Кауффман (с. 156-254)
Введение. «Массовое вымирание» в конце мела обычно рассматривается как биологическая катастрофа, заключающаяся в почти одновременном глобальном уничтожении систематически и экологически разнообразных таксонов, характерных для мезозойской биоты в целом (см., например [97]). Созданию такого представления об этом событии способствовало широко распространенное несогласие на границе мела и палеоцена, а также изображение вымирания в виде простых графиков, где линии равной толщины, представляющие главные группы ископаемых, доведены до границы эр, но не пересекают ее (например, [120], а также рис. 8.1, а). Рептилии, аммониты, рудисты, иноцерамы, склерактинии, планктонные и крупные бентосные фораминиферы, а также известковый нанопланктон—вот наиболее пострадавшие «жертвы» этого кризиса, глубоко потрясшего морские и наземные экосистемы. События конца мезозоя—начала кайнозоя, важные для понимания природы границ между эрами, являются замечательным примером биологического кризиса и находятся в фокусе многих гипотез о причинах массовых вымираний (см. [24]).
Однако «структура» этого вымирания, т. е. характер последовательности и взаимодействий биологических и экологических изменений на рубеже мела и палеогена и вблизи него, в деталях не исследована; исключением являются известковые микробиоты, изученные по данным скважин глубоководного бурения и нескольких морских разрезов на суше {Губбио в Италии, Сумайа в Испании, Тампико в Мексике и др., а также главные компоненты биоты в разрезах Дании). Этими исследованиями ясно нарисована картина глобальной катастрофы, поразившей сообщества пелагических тепловодных известковых микроорганизмов, однако при более широких обобщениях значение этого специфического события часто преувеличивается. Данные по другим группам морских организмов, особенно по макрофауне, фактически не дают достаточных оснований для «катастрофистской» интерпретации вымирания в конце мела. Большая часть тех групп, которые традиционно считаются исчезнувшими непосредственно на рубеже мела и палеогена, в действительности либо вымерла за один или несколько миллионов лет до конца Маастрихта, либо ко времени событий на этом рубеже уже испытывала эволюционный спад и была представлена только несколькими неспециализированными формами. Имеющиеся данные, взятые в целом, говорят не о всеобщей катастрофе, а о сложном характере вымирания, которое продолжалось миллионы лет.
Кроме того, концепция глобальной катастрофы, захватывающей систематически и экологически различные группы организмов, противоречит теоретическим положениям биологии, согласно которым группы с разной стратегией адаптации и различными уровнями экологической устойчивости должны (каждая по-своему) реагировать на изменения условий среды и, следовательно, их вымирание должно было происходить различным образом. Тщательные исследования эволюционных изменений на детальной стратиграфической основе действительно обнаруживают такие различия, но эти важные данные просто теряются на тех примитивных графиках, которые так часто используют катастрофисты для изображения вымирания в конце мела (в качестве примера можно сравнить различные изображения истории аммонитов на рис. 8.1, а, б). В условиях проявления реальных сил естественного отбора «тотальное» вымирание нереально как в эволюционном, так и в экологическом отношении.
Предположение о биологической катастрофе в самом конце мела не учитывает также количественную и качественную ограниченность данных, которые можно получить, изучая морские разрезы. Многие исследователи неверно интерпретируют имеющиеся факты, рассматривая их вне тех в настоящее время хорошо известных хронологических, стратиграфических, экологических и географических «рамок», которыми контролировались Маастрихтдатские события. Известно, например, что около 90% пограничных разрезов лишены слоев (и биоты) самого верхнего мела и/или самого нижнего палеоцена и из-за широко распространенного регрессивного несогласия имеют пропуск, соответствующий 2—5 млн. лет, т. е. промежутку времени, достаточному для того, чтобы вымирание приобрело сложную структуру и разные группы вымирали каждая своим темпом. В тех редких пограничных разрезах, которые обладают сравнительной полнотой, детально изучены только микроорганизмы; макрофауна обычно редка или отсутствует.
Таким образом, палеонтологическая летопись позднемаастрихтской эволюции большинства тех групп, которые считаются вымершими в конце Маастрихта, оказывается весьма неполной. Бесспорным исключением в этом отношении являются пограничные разрезы северной умеренной области в Северо-западной Европе, например в Дании. Однако именно в этих разрезах степень вымирания макрофауны и бентосной микрофауны умеренная или низкая и оно проявилось главным образом на видовом уровне; кроме того, исчезли такие группы, как аммониты и иноцерамы, которые в позднем Маастрихте находились уже на последних стадиях эволюционного спада. Весьма близкие экосистемы существовали на рубеже мела и палеогена в других частях Северо-Западной Европы, на северо-востоке США и, вероятно, в Гренландии, т. е. в Атлантической провинции северной умеренной области. В Антарктике то же самое наблюдалось Цинмейстером (личное сообщение, 1979). Во всех этих областях только сообщества известковых пелагических микроорганизмов фиксируют катастрофические события в океане; степень вымирания неизвесткового микропланктона в тех же разрезах от умеренной до низкой (Hansen, 1979, с. 141). В противоположность этому бентосная тропическая шельфовая биота тетической области (особенно организмы-рифообразователи) действительно была уничтожена в среднем и позднем Маастрихте. При этом многие быстро и повсеместно вымершие тропические группы находились вблизи пика всего эволюционного развития. Различия в характере терминального мелового вымирания в тропической и умеренной областях очень важны, но до сих пор они не получили интерпретации. Объяснение того, что случилось в тропическом море Тетис и на его тепловодных окраинах, может оказаться ключом к тайне вымирания, происшедшего в самом конце мела.
Биологическая структура терминального мелового вымирания в значительной степени остается неизвестной, хотя это событие служит классическим примером крупнейшей катастрофы. Из-за явной противоречивости существующих данных возникает много вопросов. Какова детальная последовательность событий, приведших к вымиранию? Как выяснить природу вымирания при столь неполной палеонтологической летописи события? Происходили ли в меловом периоде другие подобные события?
Если происходили, то не может ли их изучение пролить свет на природу терминального вымирания? Как можно объяснить высокую степень вымирания в тропиках по сравнению с более слабыми пограничными изменениями в северном и южном умеренных поясах? Как можно связать внезапное исчезновение многих элементов известкового микропланктона и бедность разрезов ископаемой макрофауной с выявляемым в этих же разрезах более постепенным ее вымиранием еще до конца мела? Какую из многих теоретических причин массового вымирания: быстрые температурные изменения, уменьшение солености, всеобщую регрессию, сужение экологического пространства, мощную внеземную вспышку радиации, метеоритный удар, изменение состава воды или атмосферы и т. д.—можно привлечь отдельно или в комбинации с другими для лучшего объяснения этого явления?
Данное исследование является попыткой ответить на некоторые из этих вопросов в свете современных эволюционной и экологической теорий. Для этого на соответствующих разрезах детально проанализирована «запись» последовательности событий при вымирании и зафиксированы пропуски в этой записи. Поскольку известно слишком мало полных морских разрезов перехода от мела к палеогену, в особенности разрезов, содержащих ископаемую макрофауну, ответы на вопросы, связанные с терминальным меловым вымиранием, искались в хорошо сохранившихся разрезах крупных внутримеловых вымираний. Наиболее драматичные из них приурочены к рубежам альба и сеномана, сеномана и турона, а также турона и коньяка. Биологическая структура этих, пусть менее тяжелых, но все же важных, вымираний, как правило, обнаруживает некоторые существенные особенности, которые позволяют по-новому интерпретировать терминальное меловое вымирание.
События на границе мела и палеогена: обзор представлений. Шиндевольф [119, 120] представил вымирание в конце мела как практически одновременное уничтожение многочисленных экологически разнообразных крупных групп мезозойских организмов (рис. 8.1,а); на основе этого представления им была разработана концепция «неокатастрофизма». Ньюэлл [98] рассматривал границу мела и палеогена как «массовое вымирание» и как «биологическую революцию». Персивал и Фишер [104] назвали это событие «кризисом биоты». Очевидно, что предложенная еще Кювье (1769—1862) и д'Орбиньи (1802—1857) концепция катастрофизма и массовых вымираний жива и сейчас, хотя тщательные научные исследования привели к некоторой ее модификации. В частности, идея внезапного массового вымирания на рубеже мела и палеогена глубоко укоренилась в современной науке.
Такая концепция не лишена оснований. Почти везде на границу мела и палеогена действительно приходятся резкие биологические изменения, которыми охвачены многочисленные и экологически разнообразные группы морских и наземных организмов; многие типичные меловые таксоны полностью исчезают в Маастрихте. Однако эффект этих изменений был усилен повсеместным регрессивным несогласием, приведшим в большинстве пограничных разрезов к перерыву, соответствующему от 1 до 5 млн. лет (см. рис. 8.4).
Но и в непрерывных пелагических переходных разрезах, таких, как Сумайа, в Испании, и Губбио, в Италии, на этой границе хорошо задокументировано внезапное вымирание пелагических известковых микроорганизмов на интервале около 1 м (например, [104, 105], статьи в [24, 10]). Приуроченность к маастрихтскому веку (70,5—65,6 млн. лет назад) вымирания как наземных, так и морских животных, как бентосных, так и пелагических сообществ, как простых, так и очень сложных организмов ясно указывает, что на конец мела приходится глубокий кризис биоты. Я не оспариваю реальности этого события, но, базируясь на анализе существующих палеобиологических и палеоэкологических данных, хотел бы оценить степень его внезапности, рассмотреть его «структуру», т. е. природу его биологических и экологических особенностей, и установить его причины.
Представления о катастрофическом терминальном меловом вымирании были выработаны без учета крупных стратиграфических пропусков и без детального изучения макрофауны на границе мела и палеогена, т. е. в значительной мере фактически при отсутствии данных. К тому же неверная концепция мелового вымирания исторически сложилась под определенным субъективным влиянием. Варианты таких искаженных представлений показаны на рис. 8.1—8.3.
Более ранние обзоры событий конца мела, например [119, 120, 97], в целом рисуют нам картину вымирания типичных мезозойских биот на уровне семейств, надсемейств и отрядов, не разделяя те таксоны, которые исчезли на пике эволюции, и те, которые были представлены последними эволюционными ветвями. Однако не могут считаться равноценными сравнительно быстрое вымирание в Маастрихте таких рудистов, как радиолитиды и гиппуритиды, испытывавших в это время максимум радиации (рис. 8.2,6), и окончательное вымирание столь же важного мелового семейства иноцерамов, которое к концу Маастрихта было представлено только двумя родами, несколькими видами и редкими экземплярами. События в этих двух группах существенно различны для выработки теории вымирания и понимания его причин; тем не менее многие авторы ссылаются на них как на одинаково важные компоненты терминального мелового «кризиса» (например, [104]). Серьезные интерпретации должны включать анализ таксонов низких порядков и деталей их хронологического и стратиграфического распространения как ниже, так и выше уровня вымирания (например, рис. 8.1,3 и 8.2, о, а также [104]).
Тот способ, при котором эволюция таксонов, особенно высших, графически изображается в виде постоянных по толщине прямых линий, проведенных от точки возникновения до точки вымирания (рис. 8.1, а, в и 8.2,6), также порождает неверные интерпретации природы вымираний. Такая графическая техника маскирует динамику видообразования, колебания в разнообразии таксонов, эволюционные спады и претерминальные вымирания, т. е. те данные, от которых должна зависеть интерпретация любого вымирания. Уже использованный пример рудистов и иноцерамов подходит и в данном случае. Такова же история аммонитов: изображение Шиндевольфом аммонитовых надсемейств в виде прямых линий (рис. 8.1, а) подразумевает, что их массовое вымирание произошло в конце мела. Однако графики родового разнообразия (рис. 8.1,6 и 8.5) показывают, что аммониты достигли эволюционного пика и испытали наибольшее вымирание уже в середине мела; в позднем Маастрихте сохраняются очень немногие роды и виды (см. [10]), и их окончательное вымирание не было драматичным.
Нехватка детальных биостратиграфических данных по вымершим в Маастрихте организмам также препятствует всестороннему анализу терминального мелового вымирания. За существенным исключением исследований тепловодного известкового планктона (см. [58, 104]) и макрофауны в разрезах Северо-Западной Европы (статьи и ссылки в [24, 10]), большая часть опубликованных работ по маастрихтским морским биотам дает, мягко говоря, лишь обобщенную картину. На графиках, используемых при обсуждении вымирания (например, рис. 8.1- 8.3), стратиграфический диапазон таксонов в большинстве случаев наносится с точностью не более чем до подъяруса, а в некоторых случаях только до яруса. Следовательно, какова бы ни была эволюция таксона, если он присутствует в основании Маастрихта, точка его вымирания автоматически наносится на границу мела и палеогена. Однако Маастрихт продолжался 5 млн. лет [67, 138). Этого времени было достаточно для крупных эволюционных изменений, если учесть быстрые темпы эволюции и вымирания разнообразных в систематическом и экологическом отношениях морских меловых организмов [62, 63, 69, 71]. К тому же в течение 5 млн. лет эпизоды вымирания могли как группироваться (что указывало бы на катастрофическое событие), так и распределяться случайным образом. Такие графики, как рис. 8.1—8.3, мало говорят нам о терминальном вымирании и создают искусственный образ биологической катастрофы, доказательств которой не приводится. Графики такого типа резко отличаются от тех прекрасных данных, которыми доказывается массовое вымирание известковых планктонных микроорганизмов в конце Маастрихта (например, [104], статьи в [24, 10]). Подобных данных нет по большинству тех макрофаунистических эволюционных групп, для которых также предполагается внезапное вымирание в это время.
Обращаясь к терминальному меловому вымиранию морских организмов, многие авторы не сумели заметить, что во всех обнаженных разрезах крупное параллельное или скрытое несогласие уничтожило данные о миллионах лет биологической истории в зоне перехода от мела к палеогену [98]. Это несогласие отражает размыв во время крупного эвстатического понижения уровня моря в Маастрихте и отступания береговой линии примерно на 100 м ниже ее современного положения [74, 75] (Hancock and Kauffman, 1979). В платформенных областях пропуск обычно увеличивается из-за того, что вблизи контакта развиты континентальные толщи. Если же в платформенных разрезах все-таки есть морские слои Маастрихта и палеоцена, то по крайней мере верхний Маастрихт и нижний палеоцен в морских фациях обычно отсутствуют (рис. 8.4). Этот пропуск, соответствующий 3—2 млн. лет и более, достаточно продолжителен для развития значительных эволюционных изменений и различных по стилю вымираний; поэтому чем больше перерыв, тем резче выражены различия между кампан-маастрихтскими и палеогеновыми биотами по обе стороны от него. Графики, приведенные на рис. 8.1—8.3, не показывают диастемы на этой границе; тем самым они дают неправильное представление о характере вымирания.
Попытка представить в обобщенном виде длительность перерывов в маастрихтских морских разрезах Евро-Американского региона и некоторых других мест показана на рис. 8.4. Во внутренних частях материков, например таких, как Центральная Европа, центральная часть Южной Европы и основная часть Западного Внутреннего бассейна США, морские породы Маастрихта и палеоцена, как правило, не сохранились или в лучшем случае сохранились только породы нижнего и нижней части среднего Маастрихта, перекрытые неморскими меловыми—палеоценовыми слоями, которые разделяются одним или несколькими несогласиями. Они в свою очередь перекрываются сокращенным разрезом нижнего—среднего палеоцена, представленным чередованием морских и лагунных солоноватоводных фаций (например, североамериканский разрез: морская формация Фокс-Хиллс, континентальные отложения Хелл-Крик и нижняя часть формации Ланс, морские—Кейнонбол). В наиболее полных американских платформенных разрезах между морскими фаунами есть пропуск, соответствующий времени 2—3 млн. лет.
Такие же, но, очевидно, менее значительные пропуски существуют и в разрезах континентального шельфа. В пределах равнин, обрамляющих Мексиканский залив и Атлантическое побережье, слои, содержащие богатую среднедатскую морскую фауну, лежат с параллельным или скрытым несогласием на слоях с морской фауной среднего Маастрихта. Хотя в некоторых случаях контакт проходит внутри глауконитовых песков без особых признаков эрозионного перерыва (например, в Мэриленде, где датская формация Брайтсит залегает на маастрихтской формации «Монмаут»), микропалеонтологические данные показывают, что наиболее молодые меловые породы относятся к началу среднего Маастрихта (зона Globotruncanagansseri), а перекрывающие их палеоценовые породы относятся уже к среднему данию (зона Globigerina doubjergensis—Globorotalia pseudo-bulloides). Тем самым устанавливается уменьшающийся в сторону океана пропуск минимальной продолжительностью 2,5— 3 млн. лет. Разрезы европейского шельфа аналогичны; исключением являются районы активной тектоники окраин Средиземного и Северного морей. В этих районах опускание краев континента и тектонических бассейнов привело к накоплению почти полного разреза в пелагических фациях и фациях внешнего шельфа. В Южной Америке и Южной Африке развиты преимущественно неполные пограничные разрезы мела и палеогена, которые, по-видимому (с учетом слабой изученности), подобны разрезам северной умеренной области. Центральные, тетические, области, такие, как Антиллы, Центральная Америка и окраины Карибского моря, были тектонически активными участками с развитием островных дуг; тем не менее в отложениях карбонатных платформ этого региона маастрихтские, а в большинстве случаев и палеоценовые слои отсутствуют. С другой стороны, пелагические глубоководные фации в районе Тампико, Мексика, в северо-восточной Бразилии и на Гаити образуют довольно полные разрезы, однако почти не содержащие сохранившейся макрофауны.
Таким образом, почти везде крупные стратиграфические пропуски характерны для эпиконтинентальных разрезов на переходе от мела к палеогену. Лишь в немногих обнаженных пелагических разрезах и в кернах глубоководных скважин сохранились переходные слои. Но в большинстве случаев эти разрезы и керны позволяют анализировать только микробиоту, потому что макрофауна (за исключением ходов илоедов) в таких фациях сохраняется крайне редко. Заметным исключением являются разрезы писчего мела в Скандинавии. Поэтому сохранилось очень мало настоящих доказательств того массового вымирания крупных морских организмов, которое, согласно общему мнению, произошло в самом конце позднего мела. В противоположность этому великолепные данные подтверждают внезапное катастрофическое вымирание тепловодного известкового микропланктона во время этого терминального «события».
Основные перспективы описания терминального мелового вымирания морской биоты как целого связаны с определением и детальным изучением всех групп ископаемых организмов в тех обнаженных разрезах эпиконтинентальных бассейнов, континентальных шельфов и склонов, которые накапливались над уровнем компенсации арагонита (УКА) и в которых в значительной степени сохранилась непрерывная последовательность морских осадков в интервале от верхнего Маастрихта до дания. Таких разрезов немного; в большинстве из них хорошо изучены микроорганизмы, но о микрофауне, если исключить самые мелководные и самые северные из них разрезы Северо-Западной Европы, пока еще известно слишком мало. Однако даже общий обзор этих разрезов дает новую информацию, относящуюся к биологической структуре вымирания в самом конце мела.
Характер позднемелового вымирания морских организмов: общий обзор. Концепция катастрофического терминального мелового вымирания, охватившего морфологически и экологически разнообразные группы организмов, не подкреплена достаточным количеством детальных филогенетических исследований и не вытекает из современных экологических и эволюционных теорий, согласно которым даже крупные вымирания должны быть постепенными. В предельном случае небольшая изолированная популяция стенобионтных организмов, занимающих очень узкую экологическую нишу и ограниченно распространенных, может быстро эволюционировать, а под влиянием даже слабого стресса внезапно вымирать. Другим предельным случаем являются неспециализированные организмы, существующие в широком экологическом диапазоне, образующие крупные и распространенные популяции, которые в тех же условиях могут эволюционировать медленно, а вымирать долго и постепенно. Между этими двумя типами могут быть все переходы, и такие соотношения остаются в силе на любом таксономическом уровне. Теоретически катастрофическое вымирание возможно только в результате глобального, быстро развивающегося кризиса условий обитания, изменения которых превышают уровень устойчивости некоторых эврибионтных групп, таких, как последние иноцерамы, и, уж само собой, превышают уровень устойчивости большинства сосуществующих с ними стенобионтов. Такая катастрофа может объясняться либо вмешательством внеземных сил, таких, как взрывы сверхновых или всплески радиации, превышающие обычный для Земли уровень, либо быстрыми и почти одновременными крупномасштабными изменениями ряда параметров среды, которые, действуя совместно, превышают пределы, допустимые для выживания экологически различных групп. Оба сценария предлагались для объяснения катастрофического вымирания разнообразных групп в конце мела. Однако детальные исследования характера вымирания большинства таксонов и ухудшения условий их обитания в самом конце мела показывают, что эти процессы были растянуты на миллионы лет, и, таким образом, не подтверждают концепции единственного внезапного глобального кризиса биоты.
Многие крупные изменения среды обитания действительно совпадают с концом мела: эвстатнческое понижение уровня моря примерно на 100 м ниже его современного положения, глобальная регрессия в эпиконтинентальных бассейнах, которая привела к резкому сокращению морского экологического пространства и уничтожению важных экологических ниш, быстрые флуктуации температуры морской воды в некоторых регионах, а также перестройка океанической циркуляции, которая, вероятно, сопровождалась повсеместными изменениями химического состава воды (содержания кислорода, солей, микроэлементов и т. д.). Но эти события были также растянуты во времени, по крайней мере на последние несколько миллионов лет мелового периода. Финальное эвстатическое понижение уровня океана и регрессия начались в позднем кампане и быстро развивались после эпизодической трансгрессии в среднем Маастрихте (см. [74]). Флуктуации температуры морской воды, приведшие к общему ухудшению условий, начались в среднем Маастрихте. Экологическая теория предсказывает к тому же, что вымирание, к которому привели эти изменения, должно было по-разному воздействовать на группы с различными экологическими стратегиями и устойчивостью. Некоторые из них должны были остаться почти незатронутыми, на другие должен был повлиять только наиболее мощный стресс в самом конце мела, третьи должны были исчезнуть задолго до конца мела или из-за небольших изменений, связанных с началом терминальной регрессии, или из-за еще более ранних и менее интенсивных изменений океанических условий и экологических структур. Внезапное исчезновение некоторых групп (например, известковых микроорганизмов), которые повсеместно вымерли на рубеже мела и палеогена или очень близко от него, обусловлено скорее мощностью и разнообразием совпавших во времени ухудшающих среду факторов, нежели с любым из этих факторов в отдельности или со скоростью такого ухудшения.
К тому же характер тех изменений среды обитания, которые были приурочены к концу мела и которые рассматриваются как причина терминального мелового вымирания, нельзя считать уникальным: подобные изменения меньшего масштаба характерны и для других частей мелового периода. Например, конец альба ознаменовался крупной глобальной регрессией, падением температуры морской воды (Kauffman, 1977) и бескислородным событием в океанах [43]; сочетание этих изменений привело к повсеместному усилению вымирания. Переходы от турона к коньяку и от сеномана к турону характеризуются относительно быстрым подъемом температур в эпиконтинентальных морях и/или ростом солености, вслед за чем сначала следовало широкое распространение бескислородных условий во время максимума трансгрессии и глобального звстатического подъема, а затем падение температуры во время регрессии. Границы между этими ярусами в значительной мере подчеркнуты серией близких по времени относительно быстрых вымираний, которые отражали сильный стресс, связанный с многократными изменениями условий обитания за короткий промежуток времени. Внутримеловые кризисы оказали более глубокое воздействие на вымирание многих морских групп, например аммонитов, чем терминальные события. В других группах события, связанные с терминальным меловым ухудшением условий, оказались более глубокими, но все-таки похожими на события, несколько раз происходившие в течение мела. Поэтому внутримеловые вымирания, данные о биолого-экологических особенностях которых так хорошо сохранились во многих регионах, могут детально изучаться в качестве природных моделей для интерпретации хуже сохранившейся записи событий терминального мелового вымирания.
При этой интерпретации важно учесть еще один дополнительный фактор. Вымирание группы во время или вблизи пика ее разнообразия действительно означает биологическую катастрофу (как это случилось, например, с радиолитидами и гиппуритидами, кокколитофоридами и планктонными фораминиферами; см. рис. 8.1,3 и 8.2,6). Вымирание групп вблизи их эволюционного спада, когда они представлены только несколькими неспециализированными таксонами (как это было, например, с иноцерамами; см. рис. 8.1,6), вероятно, мало что значит даже в тех случаях, когда вымирание этой группы совпадает с более драматическими событиями (см. рис. 8.2,6). Уничтожение эволюционно «старых» групп во время массового вымирания скорее всего вызывается биологическим кризисом (например, разрушением экологической структуры) и не является непосредственным отражением или тем более следствием первичных изменений в условиях обитания. Важность этих соображений иллюстрируется рис. 8.1,6, 8.2,6 и 8.5—8.9, на которых изображено развитие отдельных групп меловых моллюсков, находившихся к моменту терминального мелового вымирания на разных стадиях своей эволюции.
Аммониты часто упоминаются как крупная группа, внезапно вымершая в конце мела. Неокатастрофистскому взгляду на аммоноидей (рис. 8.1,а; [120]) как на важный компонент меловых фаун вплоть до их «внезапного» терминального вымирания способствовали графики в виде прямых линий одинаковой толщины, такие, как рис. 8.1, а, а также отсутствие стратиграфической основы, более детальной чем ярусная шкала. Рис. 8.1, б изображает это же событие по-другому: здесь на ярусной шкале, охватывающей время от девона до мела, показано родовое разнообразие надсемейств. В результате этот график ясно показывает, что максимального обилия и разнообразия аммониты достигали в период от триаса до середины мела; он показывает также, что во время их якобы внезапного вымирания в позднем Маастрихте они были представлены только несколькими находящимися на спаде филогенетическими группами. Такое представление данных несколько снижает эффект так называемого вымирания. На рис. 8.5 на подъярусной шкале более точно изображены количественные показатели возникновения, разнообразия и вымирания меловых аммонитовых родов. График показывает, что эволюционные изменения и вымирания этих родов были эпизодическими, кроме того, он указывает на связь этих эпизодов с глобальными эвстатическими флуктуациями, порождающими трансгрессивно-регрессивные циклы. Шесть из восьми главных вспышек видообразования коррелируются с трансгрессиями, потому что это были периоды расширения экологического пространства, возникновения новых экологических ниш, улучшения климата и ослабления всех видов стресса, кроме конкуренции и давления хищников. Все значительные вымирания происходили между пиком трансгрессии и пиком регрессии. Регрессии были временем увеличения стресса, связанного с сокращением экологического пространства, потерей привычных мест обитания, падением температур и повсеместным ухудшением среды. Терминальное меловое вымирание аммонитов соответствует этой модели—крупная регрессия, уменьшение содержания кислорода в океанах и быстрые температурные изменения происходили именно в это время. Самый важный аспект рис. 8.5—это демонстрация того, что все внутримеловые вымирания, кроме одного поздневаланжинского, были более значительными, чем вымирание в конце мела, и, по-видимому, столь же внезапными. Тщательное изучение крупнейшего мелового вымирания аммонитов, происшедшего в конце альба, и исследование его причин могут пролить свет на вымирание в Маастрихте.
Наконец, в тех районах, где разрез слоев, переходных от верхнего мела к палеоцену, достаточно непрерывен, можно получить некоторые очень подробные данные о природе вымирания аммонитов в течение Маастрихта. Видманн [142] описал постепенное исчезновение аммонитовых родов и видов в кампан-маастрихтском разрезе Испании, где в Сумайе и других местах сохранился полный переход от мела к палеогену в морских пелагических отложениях. Его данные, по-видимому, указывают, что морфологически более специализированные и/или географически более ограниченные таксоны (стенобионты?) во время этого финального эволюционного спада вымирают раньше, а менее специализированные и более широко распространенные роды (например, пахидисциды) исчезают позднее, в течение Маастрихта. Особенно показательна история исчезновения аммонитов, изученная Видманном в разрезе Сумайи (устное сообщение, 1977): географически более ограниченные тепловодные формы (стенобионты?) исчезают вблизи границы среднего и верхнего Маастрихта; умеренно разнообразные космополитные аммониты (эврибионты?) встречаются в нижней части верхнего Маастрихта, а затем постепенно начинают исчезать; несколько последних групп, представленных только неспециализированными космополитами, мельчают (карликовость?) и постепенно исчезают на протяжении последних 75 м разреза. Непосредственно под границей палеогена остается только один космополитный вид пахидискусов. Эти данные означают, что позднемеловое вымирание аммонитов в тетической области было постепенным: первыми исчезали экологически специализированные виды, а последними - неспециализированные. Если этот разрез представителен, то постепенное вымирание пелагических тетических аммонитов, вероятно, было связано с изменениями температуры и солености бассейна и с усилением давления хищных рыб и рептилий в течение кампана и Маастрихта, а не с какими-либо экологическими бедствиями в самом конце мела.
Рассмотрение разнообразных групп ископаемых, которые испытали заметное вымирание в конце мела, позволяет выделить четыре главных типа этого процесса (на рис. 8.6—8.9 они иллюстрированы примерами из различных семейств двустворок). Тип вымирания каждой группы часто зависит от ее эволюционной «стадии» или древности и, следовательно, от того, насколько она испытала действие конкуренции, давления новых хищников, потери основных ареалов и т. п. в течение всего мела; отчасти этот тип зависит и от того, насколько устойчива данная группа к характерным для мела резким эпизодическим изменениям условий обитания. Из этих четырех типов только один представляет драматические, т. е. действительно катастрофические, маастрихтские вымирания разветвленных групп во время пика их радиаций, причем вымирания этого типа охватили менее чем 25% групп, исчезнувших в самом конце мела. Значительно более сложные примеры длительного, постепенного и/или слабого мелового вымирания, объединенные в другие три типа, имеют совсем иное объяснение. К ним относится подавляющее большинство вымерших в конце мела таксонов, и, таким образом, именно эти три типа определяют нормальную картину вымирания.
1-й тип вымирания характерен для геологически древних эволюционных групп, которые возникли и прошли пик эволюции до мезозоя, а в мезозое испытывали эволюционный спад. Этот спад объяснялся главным образом конкурентным вытеснением и замещением со стороны более развитых организмов. В меловое время эти группы достигли последних стадий своего эволюционного спада и были представлены лишь несколькими неспециализированными таксонами, популяции которых, как правило, были немногочисленны и в некоторых случаях ограничены по ареалу, который соответствовал последнему «экологическому убежищу». Многие из этих групп вымерли на рубеже мела и палеогена или вблизи него, но их окончательное вымирание относилось к одному или, самое большее, к нескольким видам и, по-видимому, было связано скорее с общим ухудшением морских экосистем (например, с изменением структуры пищевых цепей), чем с первичными физическими причинами этого ухудшения.
Среди двустворок этот тип характерен для семейства Permoforidae, расцвет которого относится к палеозою (рис. 8.6). Во время мезозойского этапа интенсивного видообразования среди зарывающихся двустворок эта группа примитивных, полузарывющихся и неглубоко зарывающихся, бессифонных цедильщиков была вытеснена более передовыми, имеющими сифон формами [129]. Мезозойские реликтовые виды этой группы встречаются главным образом как эндемики в географически разобщенных центрах, причем некоторые в опресненных окраинных бассейнах, которые могли являться временными убежищами для этой морской группы. В течение позднего мела новые адаптивные формы, возникшие в этих биотопах, в конце концов вытеснили последних пермофорид; отдельные редкие миоконхины доживают до Маастрихта, но ни одна из них не встречена в верхнем Маастрихте. Вымирание пермофорид связано главным образом с конкурентным вытеснением; хотя изменения условий обитания, связанные с терминальной меловой регрессией, и охватили окраинно-морские биотопы, населенные реликтовыми пермофоридами, они не могут рассматриваться как первопричина исчезновения этой группы.
2-й тип вымирания характерен для групп, которые возникли в конце юры или начале мела, достигли максимального разнообразия в среднемеловую эпоху и были на стадии эволюционного спада во время маастрихтского вымирания. Маастрихтские представители некоторых из них были еще довольно разнообразны, и вымирание было значительным, однако в большинстве случаев эти группы в Маастрихте были представлены только несколькими неспециализированными видами. Этот тип вымирания характерен для меловых аммонитов (рис. 8.1,6 и 8.5), ипоцерамов и рудистов из семейства Requieniidae (рис. 8.7). Крупный спад в эволюции всех этих групп в конце мела может быть объяснен главным образом экологическими факторами: давлением хищников, изменениями в биотопах и конкурентным вытеснением. Так, в течение верхнего мела возрастало давление хищников на аммонитов; это обстоятельство вместе с общим ухудшением среды в наиболее благоприятных для аммонитов биотопах (толще воды и придонных ее слоях), вероятно, вызвало их эволюционный спад. Окончательное вымирание аммонитов, по-видимому, связано с заметными изменениями состава и температуры воды в Маастрихте. Иноцерамы, обитавшие преимущественно на поверхности дна, в значительной степени стали, жертвами крабов и питающихся донными животными позвоночных, разнообразие которых возрастало в течение позднего мела.
После возникновения в поздней юре рудисты семейства Requieniidae (рис. 8.7) быстро проникли на рифогенные карбонатные платформы и, распространившись там, стали важным компонентом рудистовых построек «среднего» мела [81]. Но их лежачие или полулежачие спиральные раковины были не лучшим образом приспособлены для строительства каркаса, и в верхнемеловых рифах они в значительной степени замещаются более вертикальными и быстрорастущими, эффективно группирующимися гиппуритидами и радиолитидами. Этот процесс привел к устойчивому спаду реквиениид в течение позднего мела (рис. 8.7); только одна неспециализированная группа (Bayleia), вероятно, сохраняется после раннего Маастрихта. Но даже если это и так, палеонтологическая летопись этих и многих других меловых рудистов не продолжается в течение последних 1 или 1,5 млн. лет маастрихтского века. Таким образом, вымирание реквиениид и других примитивных рудистов в Маастрихте (рис. 8.2, б) связано прежде всего с конкурентным вытеснением более продвинутыми рудистами и произошло до «кризиса» биоты и условий обитания в конце Маастрихта. Только внезапное исчезновение радиолитид, гиппуритид и некоторых капринид в среднем и начале позднего Маастрихта действительно имеет отношение к проблеме терминального мелового вымирания и его причин.
3-й тип вымирания. К нему относится, по всей видимости, внезапное и повсеместное исчезновение разнообразных организмов во время маастрихтского пика их радиации. Такой характер вымирания обычно предполагается для большинства таксонов, не переживших терминального мелового биотического кризиса, включая такие группы, как аммониты и иноцерамы, которые на самом деле испытывали в позднемеловое время эволюционный спад и в Маастрихте были представлены только несколькими неспециализированными таксонами. Меловые группы морских организмов, для которых действительно было характерно вымирание третьего типа, включают большую часть планктонных фораминифер, известкового нанопланктона, крупных бентосных фораминифер, рифостроящих склерактиний, многих устриц, тригоний, наиболее специализированных рудистов, а среди гастропод — неринеид и актеонеллид. В подавляющем большинстве это были стенотермные обитатели мелководных шельфов и верхней части неритовой области в тетической провинции с ее тропически-субтропическим климатом. Выяснение первичных причин терминального мелового вымирания связано с изучением именно этих групп.
Вымирание этого типа было, в частности, характерно для рудистов из семейства Radiolitidae (рис. 8.8). Эта группа возникла в раннем мелу, а в позднем достигла разнообразия благодаря нескольким вспышкам видообразования, которые были связаны с эвстатическими подъемами уровня моря и эпиконтинентальными трансгрессиями. Первый из четырех таких эпизодов приходится на самое начало альбской трансгрессии, два следующих совпадают с пиками сеноманской и коньяк-сантонской трансгрессий, а четвертый последовал сразу за максимумом трансгрессии в позднем кампане (рис. 8.8). При этом радиолитиды становятся важным компонентом многих типов рудистовых построек почти на всех стадиях их формирования [81]. Но в Маастрихте, достигнув максимума разнообразия, это семейство очень быстро вымирает (рис. 8.2,6 и 8.8). Точная датировка этого события—единственного крупного вымирания в истории радиолитид—неясна потому, что верхнемаастрихтские карбонатные шельфовые осадки редко сохраняются из-за повсеместной регрессии в самом конце мела. В тех карибских и средиземноморских разрезах, где, как, например, на Ямайке, наиболее молодые из среднемаастрихтских отложений представлены карбонатными шельфовыми фациями, как будто выявляется повсеместное вымирание разнообразных рудистов (в том числе и большинства радиолитид) уже в самом конце среднего Маастрихта вблизи его границы с поздним. Некоторые рудисты, и в их числе наименее специализированные радиолитиды, до позднего Маастрихта доживают, но, по-видимому, почти все исчезают до терминального вымирания. Исчезновение большинства рудистов в конце среднего Маастрихта может быть первым следствием повсеместного ухудшения условий обитания, которое привело к кризису в самом конце мела; это исчезновение в целом совпадает с первой стадией регрессии, с уменьшением содержания кислорода в океане, с быстрыми флуктуациями температуры и региональными температурными минимумами, приуроченными к концу Маастрихта.
Из четырех обсуждаемых типов вымирания, вероятно, только третий непосредственно отражает первичные причины терминального мелового кризиса биоты. К таким причинам, несомненно, относятся падение температуры в океанах и эпиконтинентальных морях в позднем Маастрихте и влияние этих температурных изменений на стенотермные группы, населявшие Тетис. Из этих групп более всего пострадали, по-видимому, обитатели верхней части толщи тропических вод—мелководный бентос и лланктонно-нектонная биота как постоянная, так и временная, подобная планктонным личинкам шельфового бентоса. Подавляющее большинство бентосных шельфовых беспозвоночных, живущих в тропиках, имеет длительно развивающиеся планктонные личинки.
4-й тип вымирания характерен для многих групп, которые были важными компонентами меловой биоты вплоть до конца Маастрихта, а в результате кризиса вымерли лишь частично (рис. 8.9). Как правило, в результате продолжающегося видообразования эти группы быстро восстанавливаются уже в палеоцене и в кайнозое становятся даже разнообразнее. Многие группы, для которых было характерно вымирание этого типа, в настоящее время представлены видами, трофически и/или экологически неспециализированными и/или предпочитающими жить на шельфах умеренной климатической зоны и в более глубоководных условиях. Тот факт, что в конце мела эти группы испытали лишь относительно слабое вымирание, скорее всего объясняется их большей устойчивостью к изменениям среды, приведшим к маастрихтскому кризису биоты; возможно также, что эти изменения нанесли более сильный удар по тропическим морским биотопам, чем по умеренным.
Типичным примером являются двустворки из семейства Те1linidae. Представители этого семейства—зарывающиеся илоеды—распространены в мелководных и глубоководных морях тропической и умеренной зон. Благодаря разнообразию зон их обитания, стабильным ресурсам пищи и общей экологической устойчивости терминальные меловые события, которые включали уменьшение содержания кислорода в морской воде, быстрые температурные флуктуации, повсеместную регрессию и разрушение планктонных сообществ в океане, должны были повлиять на них меньше, чем на стенотермный планктон или на тетических бентосных цедильщиков. Кроме того, их личинки, возможно, имели более короткую планктонную стадию и обитали в более глубоких слоях водной толщи, как это характерно для некоторых современных теллинид, поэтому они были менее чувствительны к стрессу. В меловой истории теллинид эпизоды быстрого видообразования в конце альба, сеномане и раннем кампане совпадали с трансгрессиями, которые, вероятно, отражали эвстатические подъемы уровня моря. Единственное крупное меловое вымирание в этом семействе произошло во время большой маастрихтской регрессии. Несмотря на то что теллиниды были экологически и трофически слабо специализированы, почти две трети меловых родов этого семейства вымерли в результате маастрихтского кризиса; причем вымершие роды были представлены главным образом тепловодными формами. Этот факт важен для оценки масштаба ухудшения условий обитания в самом конце мела. Палеогеновая радиация теллинид была быстрой и значительной; в конце эоцена их разнообразие превысило меловой уровень. Сейчас теллиниды находятся на очевидном пике своей эволюционной истории (рис. 8.9).
Выводы. Группы морских организмов, которые в значительной степени или полностью вымерли в конце мела, характеризуются разными историями этого вымирания, которые делятся на описанные выше четыре основных типа. Из этих четырех типов только один (третий) представлен относительно внезапным маастрихтским вымиранием групп, находящихся на пике своей эволюции, и к этому типу относится вымирание менее четверти всех таксонов, исчезнувших в конце мела. И все же именно такой тип обычно рассматривается как картина вымирания большинства групп в конце мела. Но даже когда вымирание действительно относилось к третьему типу, во всех группах (за исключением известковых планктонных микроорганизмов) его главные события происходили за миллион лет или больше до конца Маастрихта. Другие три типа вымирания отражают либо экологические факторы конкуренции и хищничества, уничтожившие ряд групп задолго до конца мела, либо относятся к наиболее эврибионтным группам, радиация которых была лишь временно прервана кризисом условий обитания в конце мела (рис. 8.9). При этом в первую очередь вымирали обитатели мелководных шельфов или верхней пелагиали в тропической и субтропической областях; это же характерно и для групп, вымирание которых относится к третьему типу. Вымирание в умеренной области было более постепенным, не столь сильным и охватило меньшее количество групп. Эти данные показывают, что позднемеловой кризис биоты был вызван изменениями среды обитания, охватившими прежде всего биотопы верхних слоев тропических вод.
Изучение структуры вымирания включает классификацию и интерпретацию индивидуальных особенностей эволюции тех групп, которые были этим вымиранием охвачены. Это очень важно потому, что позволяет выделить те вымирания, которые непосредственно вызваны главными изменениями среды обитания, и дать более ясную интерпретацию этих изменений. Так вырисовывается фон, на котором происходили другие вымирания, бывшие вторичным следствием нарушений экосистемы, и в результате становится возможной более объективная оценка терминального мелового вымирания в целом.
Сохранившаяся летопись вымирания морской макрофауны в конце мела. Лишь в немногих и разрозненных районах стратиграфические разрезы сохранили запись всей или почти всей истории морских бассейнов на переходе от позднего Маастрихта к датскому веку. Большая часть этих разрезов представлена глубоководными пелагическими отложениями, которые, если не считать фукоидов, содержат очень мало макрофоссилий. Детали стратиграфии, условий осадконакопления и биологической истории (главным образом истории микробиот) хорошо документированы только в нескольких районах, таких, как Губбио в Италии, Сумайа в Испании, Северо-Западная Европа (Голландия, Дания и Южная Швеция, Крым в СССР, Тампико в Мексике, южная часть Гаити и окрестности Ресифе в северо-восточной Бразилии). Многочисленные керны глубоководных скважин раскрывают дополнительные детали событий на границе мела и палеогена, но только шельфовые разрезы Северо-Западной Европы содержат обильную макрофауну. Однако для понимания природы терминального мелового вымирания история редкой макрофауны в пелагических разрезах открытого моря также представляет интерес. Здесь дается обзор этой истории и, кроме того, рассматриваются данные по столь же важным, но менее полным пограничным разрезам тетических карбонатных платформ (рудистовые фации).
Отсутствие или скудость раковинной макрофауны в таких глубоководных пелагических карбонатных разрезах, как Губбио, Сумайа и Тампико, связывались либо с растворением ниже уровней арагонитовой и кальцитовой компенсации, либо с общей экологической ограниченностью макробентоса в условиях глубокого моря. Ни одна из этих гипотез в целом не подкрепляется существующими данными. Характер сохранности микроорганизмов в этих же осадках исключает широкое проявление растворения в большинстве разрезов. С другой стороны, в области накопления богатых органикой глубоководных илов распространена одна из наиболее разнообразных современных бентосных макрофаун, особенно раковинных моллюсков [117]. Даже ниже уровня компенсации арагонита толстый периостракум раковин многих моллюсков защищает их от растворения некоторое время после смерти [66]. Обилие и разнообразие следов жизнедеятельности во многих пелагических осадках и в большинстве изученных пограничных разрезов опровергают предположение о том, что резкое сокращение раковинного бентоса—нормальное свойство данной фации. Малочисленность раковинных макрофаун в верхнем Маастрихте многих разрезов (например, Губбио, Сумайя и Тампико) должна рассматриваться как важное следствие терминального мелового вымирания и тех крупномасштабных экологических изменений, которыми оно было вызвано.
Тетические пелагические разрезы. Губбио, Италия. Исключительно полная последовательность тропических пелагических карбонатов заполняет Маастрихт-датский интервал в Губбио в Италии. Этот разрез был выбран в качестве стратотипа при разработке шкалы позднемеловых геомагнитных инверсий [4, 111, 89], детально изучены его литостратиграфия, седиментология (6], биостратиграфия, а также эволюционная история планктонных известковых микроорганизмов ([105], Monechi, 1977, 1979).
Контакт Маастрихта и дания, определяемый по внезапному исчезновению практически всех меловых известковых микроорганизмов, расположен внутри разреза известняков. Тонкий прослой глины залегает менее чем на 1 м выше этого контакта. Артур и Фишер [6] и Премоли-Сильва [105] приводят полный разрез всех зон планктонных микроорганизмов верхнего Маастрихта и палеоцена и не говорят о несогласии на границе между ними; но другие авторы (например, Гампер [44], ссылающийся на Лютербахера и Премоли-Сильву [89]) считают, что верхняя часть верхнемаастрихтской зоны Abathomphalus mayaroensis отсутствует, по-видимому, из-за скрытого несогласия на этой границе. Не сообщается ни о каких макрофоссилиях ни из верхнего Маастрихта, ни из самой нижней части датского яруса ([6]; в работе [105) специально подчеркивается отсутствие аммонитов). Хотя у фораминифер не обнаруживается значительного разрушения кальцита раковин в результате растворения или в процессе диагенеза, некоторое растворение кокколитов было замечено ([105], Monechi, 1979). Это означает, что если в районе Губбио на дне или в толще воды существовала макрофауна с кальцитовым или арагонитовым скелетом, то по крайней мере некоторые элементы этой фауны должны были сохраниться в разрезе.
Обильные и разнообразные следы жизнедеятельности организмов в разрезе пограничных слоев мела и палеогена в Губбио отмечаются Артуром и Фишером [6]. Согласно личному сообщению Фишера в марте 1979 г., «в кампан-палеоценовой части Скаглия-Росса первичная слоистость практически отсутствует, и хотя имеются фукоиды, известняки и мергели в основном однородны; биотурбация уничтожила детали истории их накопления».
Отсутствие бентосной раковинной макрофауны в разрезе Губбио обусловлено отнюдь не бескислородными условиями и/или обогащением субстрата или придонных вод сероводородом. В других меловых разрезах в подобных пелагических отложениях, а также в отложениях, где интенсивная биотурбация отсутствует, сохраняются такие бентосные двустворки, как иноцерамы, и другие эпи- и эндобионты: черви, усоногие раки, мшанки, устрицы, птерии и пектены (72, 73]; по-видимому, в меловое время эти группы обладали высокой устойчивостью к нехватке кислорода. Таким образом, отсутствие макрофауны в верхнем Маастрихте Губбио скорее всего отражает более раннее ухудшение условий внутри толщи морской воды; это ухудшение, по-видимому, распространилось на всю водную толщу и к позднему Маастрихту постепенно охватило и придонные биотопы. Именно это более раннее ухудшение в конце концов привело к терминальной катастрофе тепловодных верхнепелагических известковых микробиот на рубеже мела и палеогена.
Сумайа, Испания. Непрерывный пограничный разрез пелагических отложений в Сумайе, расположенный на северном побережье Испании между Бильбао и Сан-Себастьяном, содержит больше точных данных о подзнемаастрихтском вымирании, чем любой другой разрез глубоководных пелагических отложений. Персивал и Фишер [104] описали «исключительно полный осадочный разрез» глубоководного флиша, внутри которого проходит граница мела и палеогена и который включает 75 м поздне-маастрихтских «пурпурных мергелей» (зона Abathompmalus mayaroensis), которые согласно перекрыты пиритизированными «пограничными глинами» мощностью 25—35 см. Планктонные фораминиферы зоны A. mayaroensis и известковый нанопланктон проникают на 10 см вверх в пограничные глины и внезапно исчезают на границе мела и палеогена [104]. Верхние 15—28 см пограничных глин относятся уже к планктонной зоне Globigeriпа eugubina самого нижнего палеоцена. Серые палеоценовые известняки и глинистые сланцы (зона Globorotalia pseudobulloides) согласно перекрывают пограничные глины.
Структура вымирания пелагической микробиоты, отраженная в разрезе Сумайа, была описана Хермом [58] и Персивалом и Фишером [104] и сводится к следующему:
1. В нескольких метрах от контакта слои самого верхнего Маастрихта характеризуются нормальными и разнообразными сообществами океанических тетических планктонных фораминифер и известкового нанопланктона.
2. В нескольких метрах ниже кровли Маастрихта Хермом (58] отмечается уменьшение размеров планктонных фораминифер, селективное вымирание некоторых видов глоботрункан, разрушение структуры пелагического сообщества и увеличение относительной обильности известкового бентоса; морская нано-флора остается нормальной.
3. Примерно за 10 тыс. лет до терминального вымирания продолжающийся упадок планктонных фораминифер сопровождается появлением среди нанофлоры и динофлагеллят некоторых экологических неспециализированных форм, и в первую очередь браарудосфер и торакосфер (так называемые «disaster species», или виды-сигналы бедствия [104]). Появление этих форм, которые в нормальных условиях вытесняются из сложных сообществ экологически специализированных микроорганизмов, впервые фиксируется в нескольких верхних сантиметрах пурпурных мергелей и означает начало стресса и повсеместного разрушения пелагических экосистем. Во время накопления нижней половины пограничных глин виды-сигналы бедствия постепенно становятся численно доминирующими, в то время как количество нормальной нанофлоры уменьшается. Тогда же бентосные фораминиферы начинают преобладать над планктонными [58].
4. В 15 см над подошвой пограничных глин оставшиеся меловые известковые планктонные фораминиферы и нанофоссилии внезапно исчезают, вероятно, вследствие терминального вымирания.
5. В базальном палеоцене «живучие» виды браарудосфер и торакосфер становятся численно преобладающими, но в систематическом отношении остаются однообразными [104]. Планктонные фораминиферы вначале, по-видимому, были редкими и очень мелкими; вверх по разрезу они становятся более разнообразными. Этот комплекс, соответствующий по времени примерно 1 млн. лет назад, распространен в пределах 6 м, охватывающих верхнюю часть пограничных глин и нижнюю часть «датского известняка» [104].
6. За этим следует быстрое увеличение разнообразия нанофлоры и планктонных фораминифер дания и возникновение типичной палеоценовой пелагической микробиоты.
В Маастрихте Сумайи остатки макрофауны известны вплоть до первых сантиметров в основании пограничных глин; существование этих макрофоссилий дает уникальную возможность изучить вымирание бентосных и нектонных крупных беспозвоночных на материале пелагического разреза Тетической области. Видманн ([142], а также работа, готовящаяся к изданию) изучал аммониты, а Кауффман (работа готовится к изданию) — двустворки в разрезе Сумайа. Дж. Видманн (устное сообщение, 1977) любезно передал следующую характеристику распределения раковинной макрофауны в маастрихтском разрезе Сумайа: «В основании пограничных глин, в 10—15 см ниже границы мела и палеогена был найден только один мелкий (карликовый?) аммонит из группы пахидисцид. Ниже в 75-метровой пачке пурпурных мергелей изредка встречаются аммониты умеренно разнообразного космополитного комплекса, состоящего из десяти видов, принадлежащих к родам Diplofnoceras, Scaphites и Pachydiscus, все особи слишком мелкие для взрослых, по-видимому, это карликовость, появившаяся под влиянием стресса. На этом интервале разнообразие и обилие аммонитов уменьшаются вверх по разрезу. В мергелях, залегающих в интервале от 75 до 100 м и более ниже контакта, тот же комплекс представлен аммонитами с нормальным размером взрослых особей. Эти мергели подстилаются несколькими метрами не содержащих фауну известняков, а под ними, в нижней части маастрихтского разреза, залегают мощные известняки с первым раковинным макробентосом. Для этих известняков характерны две биофации: их верхние несколько метров содержат обильные иноцерамы (из эволюционного плексуса Trochoceramus radiosus, Т. somaUensis и Т. гМеИ) и явно тепловодный комплекс аммонитов; ниже такие же аммониты встречаются в ассоциации с более нормальной морской тропической бентосной фауной, которая состоит главным образом из морских ежей и моллюсков. Породы разреза Сумайа интенсивно биотурбированы почти на всех уровнях, близких к границе Маастрихта и датского яруса 132, 104]. Хотя в собственно пограничной зоне следы илоедов встречаются, они там довольно редки, что говорит о большом дефиците кислорода или сильном сероводородном заражении осадков на границе мела и палеогена».
Из данных следует такая последовательность событий в морском бассейне в районе Сумайи:
1. В конце раннего—начале среднего Маастрихта повышение океанических температур [67, 75] способствовало понижению уровня компенсации арагонита и/или кальцита и развитию нормальных морских биот на дне и в толще воды наряду с тетическими и космополитными пелагическими сообществами.
2. В конце среднего Маастрихта на фоне продолжавшегося потепления практически исчез (вероятнее всего, из-за недостатка кислорода в нижних слоях воды) самый стенобиотный тропический бентос, что позволило более эврибиотным иноцерамам заселить свободный субстрат.
3. На рубеже среднего и позднего Маастрихта исчезли жившие в толще воды тетические аммониты и весь бентос, за исключением мягких илоедов, что было связано с подъемом придонной и/или внутриводной зоны кислородного минимума в результате эвстатической трансгрессии и высоких океанических температур.
4. Начало охлаждения в позднем Маастрихте (например, [34]) привело к вторжению космополитных аммонитов из умеренной климатической зоны.
5. В самом конце Маастрихта воздействие экологического стресса на всю морскую биоту было связано с всеобщим ухудшением условий среды в результате регрессии, дестабилизации морских температур, увеличения градиента поверхностных и донных температур [11] и с расширением бескислородных зон.. Этот экологический стресс вызвал задержки в росте аммонитов и постепенное уменьшение их разнообразия, некоторое понижение плотности популяций мягких илоедов и первые изменения в ассоциациях планктонных фораминифер (58).
6. Примерно за 10 тыс. лет до окончательного вымирания известкового микропланктона исчезли последние неспециализированные формы аммонитов, уменьшилась плотность популяций бентосных роющих организмов и распространились обильные пелагические браарудосферы и торакосферы.
Западные Динариды, Югославия. Стратиграфическая и палеонтологическая последовательность в относительно полных, содержащих макрофауну пограничных разрезах Западных Динарид, представленных морскими и лагунными отложениями, в основных чертах намечена Павловецом и Пленичаром [101], но детали этой последовательности, на основании которых можно было бы выявить особенности вымирания, еще не опубликованы. По данным этих авторов в разрезе снизу вверх залегают:
1. Мелководные морские карбонаты, содержащие рудисты.
2. Очень мелководные (включающие отложения приливной зоны), не содержащие рудистов карбонаты верхнего Маастрихта («самого верхнего мела») с признаками осушения и карстообразования, соответствующими, вероятно, финальному пику регрессии (R10o по [74]).
3. Трансгрессивная серия лагунных и мелководно-морских отложений «самого нижнего палеогена».
Этот разрез напоминает разрез Маастрихта и палеоцена в Испанских Пиренеях (гарюмний), но в нем, по-видимому, преобладают прибрежные морские образования. Сведения о точном возрасте, разнообразии и экологической специализации рудистов будут очень важны для нашей гипотезы о том, что вымирание тетических рудистов произошло задолго до конца Маастрихта. Скудные югославские данные допускают такую интерпретацию.
Северо-восточная Бразилия. Мэйбисун и др. [90] описали исключительно полный разрез слоев от сантона до палеоцена между Ресифе и Жоао в северо-восточной Бразилии. Граница мела и палеогена выражена здесь «резким фаунистическим разрывом» [90] в «непрерывно накапливавшейся» последовательности микритовых пелагических известняков и кальцикластитов. Не обнаружено никаких пограничных глин. Резкие фаунистические изменения выявлены главным образом на фораминиферах, хотя сообщается о многочисленных макрофоссилиях выше и ниже границы. Этот разрез, накопившийся в теплоумеренных и субтропических условиях, выглядит как идеальное место для изучения вымирания в конце мела; к сожалению, о характере фаунистических изменений на границе, за исключением данных о фораминиферах, сообщается лишь в самом общем виде. Анализ списков фораминифер, приведенных этими авторами, указывает на существование в этом регионе крупного скрытого перерыва или слабого несогласия на границе мела и палеогена: в разрезе отсутствует как самая верхняя маастрихтская зона планктонных фораминифер Abathomphalus mayaroensis, так и самая нижняя датская зона Globigerina eugubina, в списках нет фораминифер, типичных для этих зон.
Несмотря на этот пропуск, вызывают интерес распределение макрофауны и изменения условий ее обитания как в средне-маастрихтском трансгрессивном разрезе (комплекс с Globotrunсапа coniusa, G. stuarti) ниже контакта, так и в нижнем палеоцене (зона Globorotalia pseudobulloides) над ним. Согласно описанию, это распределение выглядит следующим образом:
1. Нижняя часть среднемаастрихтской свиты Грамаме сложена мелководно-шельфовыми обломочными известняками с субтропической фауной, включающей разнообразных моллюсков, в том числе толстостворчатых двустворок, таких, как Cucul— laea и Veniella, гастропод Turritella, а также крупных ежей и обильных позвоночных.
2. Фосфатистые известняки середины среднего Маастрихта, которые накапливались вдали от берега, отвечают продолжающейся трансгрессии и содержат еще более разнообразную, хотя и более мелкую, фауну тонкостворчатых моллюсков, включая таких двустворок, как Lucina, Cardium, PUcatula, и таких гастропод, как Cypraea, Xenophora, Anchura и Turritella обильны и позвоночные. Комплекс резко обедняется в кровле этих слоев.
3. Выше, до границы мела и палеогена (где средний Маастрихт перекрывается средним данием), залегают тонкозернистые пелагические изве








