 2013-12-31
2013-12-31 1877
1877Классификация методов клонального микроразмножения.
В основе клонального микроразмножения лежит явление тотипотент-иости, т. е. способности растения структурно и функционально восстанавливаться из части, органа или отдельной клетки. Тотипотентность проявляется в клетках, называемых меристемоидами - морфогенетически компетентными клетками, которые отвечают на индукторы дифференцировки и состав сред образованием побегов, корней, зародыша. Их отличает крупное ядро, густая цитоплазма и небольшая вакуоль. Такие активно делящиеся клетки расположены в недифференцированной ткани верхушечных (апикальных) меристем, а также меристем пазушных и спящих почек стебля.
Образование меристематических тканей может быть индуцировано н культуре in vitro из специализированных тканей. В этом случае возникают добавочные меристематические ткани (процесс дедифференциации)в виде точек роста или соматических эмбриоидов. В дальнейшем происходит регенерация из меристематических тканей по пути органогенеза или соматического эмбриогенеза. Добавочные меристемы формируются непосредственно в родительской ткани, из промежуточного каллуса или клеток суспензионных культур.
В зависимости от типа меристематической ткани, используемой для регенерации, а также особенностей протекания процесса регенерации существуют три основных подхода к методам клонального микроразмножения:
1) активация развития уже существующих в растениях меристем (апекс стебля, пазушные и спящие почки стебля);
2) индукция образования новых стеблевых почек или эмбриоидов непосредственно на тканях экспланта;
3) возникновение почек или эмбриоидов из первичного и пересадочного каллуса, суспензионной культуры клеток или протопластов.
Принципиальное отличие второго и третьего подходов от первого заключается в образовании новых меристематических зон среди специализированных клеток в результате процесса дедифференциации. Первый подход является наиболее естественным и предпочтительным, поскольку в нем участвуют уже сформировавшиеся меристемы, особенностью которых является генетическая стабильность. При использовании в качестве эксплантов дифференцированных тканей растений, образующих адвентивные почки или эмбриоиды, вероятность появления мутантных форм возрастает. Особенностью третьего подхода является образование почек или эмбриоидов через стадию каллуса, суспензионной культуры или протопластов, что еще более повышает возможность возникновения мутаций.
Активация развития уже существующих в растениях меристем является основным методом клонального микроразмножения. Метод основан на снятии эффекта апикального доминирования. При удалении верхушечной точки роста в пазушных почках возрастает концентрация цитокининов, снижается концентрация ауксинов, индуцируется клеточное деление и рост пазушных почек. Снять эффект апикального доминирования также можно путем добавления в питательную среду цитокининов (6-БАП, кинетин, 2-ip, зеатин). Под воздействием цитокининов происходит также реювенилизация, т. е. омоложение тканей. Такие ткани обладают большей способностью к активному росту и регенерации побегов и корней. Эксплантами для введения в культуру in vitro при этом методе служат апикальные меристемы или кончики побегов после их стерилизации. Апикальные меристемы используются с целью оздоровления посадочного материала от вирусной, бактериальной и грибной инфекции.
Если проблема вирусов не является актуальной, целесообразно использовать кончики побегов. Следует иметь в виду, что чем больше размер экспланта, тем больше вероятность его успешной пролиферации в культуре in vitro. Однако при этом повышается и опасность инфицирования среды.
Р. Пиерик предлагает выделять метод однопочковых черенков (рис. 8.1) и метод придаточных почек (рис. 8.2).
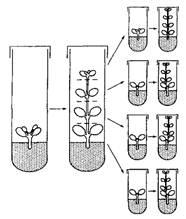 |
| Рис. 8.1 - Схема клонального микроразмножения методом однопочковых черенков |
Первый метод не требует применения цитокининов, второй основан на добавлении в состав среды цитокининов и применяется для размножения декоративных (гербера, гвоздика, розы), плодовых (яблоня, груша, вишня, слива), ягодных (земляника, смородина, малина, ежевика), лесных (береза, тополь, туя, можжевельник) растений. Второй метод клонального микроразмножения - индукция образования новых стеблевых почек или эмбриоидов непосредственно на тканях экспланта. Он включает ряд процессов: дедифференциацию специализированных клеток, клеточное деление и образование новых меристем, образование и развитие органов.
Процесс этот может происходить по пути органогенеза или соматического эмбриогенеза. Эксплант для прямого органогенеза или соматического эмбриогенеза может быть вычленен из многих органов растения (листа, стебля, зародыша, семядолей, чешуи и донца луковиц, корней, апикальных или пазушных меристем).
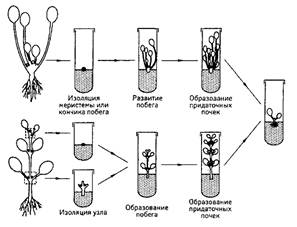 |
| Рис. 8.2 - Схема клонального микроразмножения методом придаточных почек (верхний ряд - розеточные растений, нижний ряд - удлиненные растения) |
Процесс образования адвентивных побегов лучше происходит в молодых частях растения (зародыши, проростки) или после реювенилизации тканей. Важную роль при этом играют углеводы, а гиббереллины и абсцизовая кислота его ингибируют. Процесс образования адвентивных побегов из тканей листа in vitro известен у 350 видов растений. Двудольные растения образуют почки на листьях чаще, чем однодольные.
Важным фактором является соотношение эндогенных регуляторов роста в экспланте и экзогенных - в питательной среде. Некоторые растения не требуют для образования адвентивных побегов ни ауксинов, ни цитокининов (цикорий). Для большинства растений, образующих адвентивные побеги, необходимо наличие цитокининов, которому должно предшествовать наличие ауксинов. Как правило, для таких растений необходима высокая концентрация цитокининов и низкая - ауксинов (бегония). Для отдельных растений достаточно наличия в среде только экзогенных ауксинов (лилия). В некоторых случаях возникают эмбриоиды (морковь, лен, рапс). Образование адвентивных почек на сегментах листьев и чешуи чаще всего происходит из эпидермиса.
Адвентивные побеги получены при культивировании сегментов листьев, соцветий и цветочных почек таких двудольных растений, как гербера, сенполия, бегония, маргаритка, каладиум; сегментов гипокотиля, семядолей, листьев - у представителей рода Brassica (капуста кочанная, цветная, брюссельская, листовая, брокколи); сегментов корней - у петунии.
Достигнуты успехи в клональном микроразмножении однодольных растений на основе получения адвентивных почек у представителей семейств Liliaceae, Amaryllidaceae, Iridaceae (фрезия, гиппеаструм, амариллис, гиацинт, нарцисс, гладиолусы, лилии, тюльпаны). Эксплантами в этом случае служили луковичные чешуи, сегменты донца луковицы, листья.
Третий метод клонального микроразмножения - возникновение почек или эмбриоидов из первичного и пересадочного каллуса, суспензионной культуры клеток или протопластов. Главная его особенность в том, что меристематические ткани возникают в результате непрямого органогенеза или соматического эмбриогенеза. Этому предшествует дедифференциация специализированных соматических тканей экспланта, в качестве которого могут быть использованы практически любые органы растения (корни, стебли, листья, цветки). Главная проблема, возникающая при клональном микроразмножении таким образом - возможные мутации (изменение уровня плоидности, хромосомные аберрации, генные мутации) в культуре тканей. В связи с этим существуют два основных требования при использовании такой технологии:
1) генетическая стабильность, т.е. полученный в результате размножения материал должен быть идентичен стартовому;
2) при повторном пассировании каллуса способность его к регенерации не должна быть потеряна.
Образование адвентивных побегов возможно при преобладании концентрации цитокининов над концентрацией ауксинов. Для успешного использования каллуса при клональном микроразмножении растений необходимы 2 важных условия:
а) каллус должен содержать генетически стабильные меристематические ткани;
б) каллус должен обеспечивать получение большого числа регенерантов.
Имеются сведения о получении регенерантов из каллуса у хризантем, фрезии, томатов, лилий. Второй возможный путь регенерации из дедиффе-рснцированных клеток экспланта - соматический эмбриогенез. В этом случае среди клеток каллуса возникают индуцированные эмбриогенно детерминированные клетки. Они отличаются малым размером, плотным содержанием цитоплазмы, большим ядром и малым размером вакуоли.
Условия, необходимые для успешного соматического эмбриогенеза:
1. Высокая концентрация ауксинов для индукции образования эмбриогенных клеток. Для дальнейшего развития эмбриоидов концентрация ауксинов в среде должна быть уменьшена или они должны быть полностью исключены из состава среды.
2. Гиббереллин и этилен обычно ингибируют эмбриогенез.
3. Эмбриогенезу должна предшествовать ювенилизация тканей.
4. Уменьшение содержания аммонийного азота в среде является важным фактором эмбриогенеза, наличие калия способствует эмбриогенезу, а высокая концентрация кальция ингибирует этот процесс. Образованию эмбриоидов способствует высокая концентрация солей в среде.
5. Обычно свет способствует эмбриогенезу, однако для некоторых видов этот процесс может протекать при слабой освещенности и в темноте.
6. Высокая температура обычно благоприятна для соматического эмбриогенеза; некоторые растения (особенно в культуре пыльников) требуют холодового шока для образования эмбриоидов.
7. Оптимальная концентрация сахарозы 2-3%.
Соматический эмбриогенез редко используется в практике микро-клонального размножения в связи с высокой вероятностью мутаций, трудностью метода, возможностью потери способности к регенерации при длительном пассировании каллуса. Его применяют для тех культур, где не отмечено повышение мутабильности (например, масличная пальма).
Возможно получение адвентивных побегов или эмбриоидов через стадию каллуса из одиночных клеток (суспензионная культура, культура протопластов). Получены положительные результаты в этом направлении у табака, моркови, спаржи и других культур. Проводятся исследования по разработке массового соматического эмбриогенеза с целью получения искусственных семян. Соматические эмбриоиды в отличие от семян, полученных после оплодотворения из зиготы, не имеют запаса питательных веществ. С этой целью эмбриоиды помещают в специальные полоски или капсулы с запасом питательных веществ. Возможен также посев искусственных семян в грунт с током жидкости. После разработки экономически оправданных технологий посева таким способом в перспективе можно будет размножать уникальные генотипы. Особую значимость такие технологии могут приобрести для размножения растений, не образующих семян, а также растений, у которых половое размножение затруднено (стерильные формы при получении гетерозисных гибридов).
Т. Мурасиге в 1974 г. впервые предложил подразделить микроразмножение на 3 этапа:
1. Введение экспланта в культуру in vitro.
2. Собственно микроразмножение.
3. Укоренение и адаптация в нестерильных условиях.
В последующем были разработаны схемы клонального микроразмножения, более соответствующие выполняемым задачам на каждом этапе. П. Деберг и А. Мэйн в 1981 г. предложили выделить 5 этапов (стадий) клонального микроразмножения:
Этап 0. Подготовительная стадия. На этой стадии растительный материал подготавливается к культуре in vitro. Цель этапа состоит в получении стерильного стартового материала, находящегося в оптимальном для введения в культуру физиологическом состоянии.
Этап 1. Введение в культуру in vitro. Цель - обеспечить успешную пролиферацию экспланта в культуре in vitro. Размножение на этом этапе не является важным.
Этап 2. Собственно размножение. Единственная функция этой стадии - увеличить число побегов. Для этого индуцируются меристематические центры, которые развиваются в почки и/или побеги.
Этап 3. Удлинение побегов и укоренение. На этом этапе удлиняются побеги, индуцируются и развиваются корни.
Этап 4. Перенос в тепличные условия. Большинство видов требуют адаптации в условиях ex vitro.
Рассмотрим особенности клонального микроразмножения на каждом из его этапов.
Этап 0. Подготовительная стадия.Исходным материалом для введения в культуру обычно служат элитные растения, типичные для данного сорта, без признаков инфекции. Для подготовки таких растений к введению в культуру in vitro желательно уменьшить их инфицированность путем выращивания в условиях теплиц, а также изменить физиологический статус. В теплицах поддерживается высокая температура (25°С) и сравнительно низкая влажность (70%). Для уменьшения инфицированное растения поливают непосредственно под корень, исключая полив листьев. Минимальная длительность такого режима зависит от вида растений. Например, для фикуса и драцены достаточно 3-х месяцев. На этом этапе проводится термотерапия для борьбы с вирусами.
Важно изменить физиологическое состояние растения, обеспечив реювенилизацию растений, способность тканей экспланта к пролиферации. Для этой цели регулируют физические условия (свет, температура), а также используют регуляторы роста. Целесообразно поддержание оптимального фотопериода. Важным является и спектральный состав света. Например, установлено, что длинноволновый красный свет обеспечивал повышение числа побегов у петунии в 3 раза в сравнении с необработанными растениями.
Для некоторых растений необходима холодовая обработка при температуре 4-5°С с целью выведения из состояния покоя (древесные растения). Длительность такой обработки зависит от вида растений.
Для реювенилизации исходных растений используются также регуляторы роста путем инъекции в стебель, опрыскивания или выдерживания эксплантов в специальных растворах. Для этой цели чаще всего используют цитокинины (6-БАП), а также гибберелловую кислоту.
Этап 1. Введение в культуру in vitro. Целью этапа является инициация роста тканей экспланта in vitro. Эксплантом для микроразмножения обычно служат апикальные или боковые почки. Для некоторых растений (фикус, антуриум, глоксиния) используют кусочки листа, для герберы и фрезии - части цветка. Для получения свободных от вирусов растений используют апикальные меристемы. При выборе экспланта необходимо учитывать следующее:
а) экспланты из надземной части растения менее инфицированы, чем из подземной;
б) внутренние части растения менее инфицированы, чем внешние;
в) чем меньше эксплант, тем меньше риск инфицирования;
г) регенерационная способность экспланта обычно обратно пропорциональна возрасту экспланта и возрасту исходного растения;
д) эксплант, взятый из верхушечной почки, часто находится в более молодом возрасте и регенерационная способность его выше;
е) на эффективность приживаемости и успешной пролиферации влияет время взятия экспланта. Экспланты, взятые в состоянии активно роста растения (весна - начало лета), приживаются более успешно сравнении с эксплантами, взятыми у растений, находящихся в состоянии покоя (осень - зима).
Стерилизация эксплантов приведена выше. В качестве питательной среды чаще всего используют среду Мурасиге, Скуга или ее модификации. Для древесных растений применяют среду WPM (woody plant medium).
Учитывая видовую и сортовую специфику, оптимизируют состав микро- и микроэлементов, витаминов, углеводов и регуляторов роста для каждого вида растения на каждом этапе микроразмножения. В составе питательной среды обычно используют следующие регуляторы роста: цитокинины 6-БАП, кинетин, зеатин, 2-ip в концентрации 1-10 мг/л; ауксины - индолилуксусная, индолилмасляная или нафтилуксусная кислоты в концентрации 0-1 мг/л. Для апикальных меристем наряду с цитокининами и ауксинами возможно использование гибберелловой кислоты (0-1 мг/л). Физические условия в культуральной комнате: температура 18-28°С, длина дня - 16 часов.
Важной на этом этапе, в особенности для древесных растений, является проблема полифенолов. При механической изоляции экспланта возникает стрессовая ситуация, в которой синтез полифенолов усиливается. В культуре in vitro полифенолы окисляются полифенолоксидазами. Продукты окисления ингибируют активность ферментов, вызывают потемнение ткани и среды, что может привести к гибели экспланта. Для борьбы с этим явлением можно использовать антиоксиданты путем добавления в состав питательной среды или обработки экспланта перед помещением на среду. В качестве антиоксидантов используют поливинилпирролидон, аскорбиновую кислоту, дитиотриэтол. Возможно добавление в состав среды активированного угля. Не исключены и другие подходы к решению проблемы полифенолов: культивирование эксплантов в начальный период в темноте или при пониженной освещенности, снижение температуры, частая пересадка эксплантов на свежую питательную среду. Длительность первого этапа колеблется от 1 до 2 мес.
Этап 2. Собственно размножение.Цель этапа - получение максимального количества побегов за один пассаж. Главным условием образования побегов является высокое соотношение цитокининов к ауксинов. Потребность в экзогенных гормонах зависит от концентрации цитокининов в экспланте. Среди цитокининов часто используется 6-БАП, в отдельных случаях - кинетин и 2-ip. Обычно 1-2 мг/л цитокинина достаточно для пролиферации побегов. Более высокие концентрации усиливают образование адвентивных почек, что не всегда желательно в связи с повышением вероятности образования мутаций. Возможно использование ауксинов для повышения активности роста побегов. Чаще используют нафтилуксусную и индолилмасляную кислоты в концентрации0,1-1мл/л, реже - индолилуксусную кислоту из-за ее нестабильности в среде. Ауксин 2,4-Д не применяется, поскольку может вызывать мутации.
Существуют различные способы увеличения числа побегов на этапе 2. Наиболее простой заключается в росте побега в длину в результате развития апикальной меристемы с последующим делением побега на черенки, несущие пазушные почки. Таким способом размножается картофель. Другой путь - развитие побегов из пазушных почек в результате снятия эффекта апикального доминирования под действием цитокининов. В этом случае из экспланта за 4-8 недель развивается пучок (кластер) побегов. После разделения пучка миниатюрных побегов и посадки их на свежую среду процесс образования нового пучка побегов повторяется. В обоих описанных вариантах побеги образуются из существующих меристем. Третий путь - образование побегов из адвентивных почек. Почки возникают в любом месте, кроме апикальной или пазушной меристемы. Такой путь обеспечивает более быстрое размножение, однако повышает вероятность мутаций.
Большинство коммерческих систем микроразмножения растений обеспечивает смешанный тип органогенеза, т. е. образование побегов как из пазушных, так и из адвентивных почек. Для отдельных культур возможно размножение в результате соматического эмбриогенеза.
При многократном пассировании тип органогенеза на той же среде может изменяться (вместо боковых образуются адвентивные почки). Многократное пассирование увеличивает вероятность эпигенетических изменений и мутаций. Возможно также снижение регенерационного потенциала после 7-10-го пассажа. Обычно 10-12 пассажей считаются максимумом.
Иногда на этом этапе возникает явление стекловидности или «витрификации», т.е. избыточной оводненности побегов. В таких растениях нарушается процесс образования хлорофилла, протеинов, снижается приживаемость при пересадке. Один из возможных путей уменьшения оводненности - снижение концентрации цитокининов в среде.
Этап 3. Удлинение побегов, индукция и развитие корней.Побеги, полученные на этапе 2 при высокой концентрации цитокининов, имеют небольшой размер, что затрудняет манипуляции с ними. Удлинения побегов можно достигнуть при их пересадке на среду, не содержащую цитокинины.
Пазушные или адвентивные побеги, полученные при наличии цитокининов, обычно не имеют корней. В связи с этим главная задача этапа 3 - индукция ризогенеза. Ризогенез имеет три фазы: индукция, инициация и элонгация. Первые две фазы трудно различимы. Образование корней происходит при низком содержании или отсутствии цитокининов и высоком содержании в среде ауксинов. Обычно остаточного количества цитокининов в побегах после этапа 2 достаточно и их добавления в среду не требуется. На этапе 3 чаще всего применяют нафтилуксусную, индолилмасляную, индолилуксусную кислоты в концентрации 0,1-1 мг/л.
Высокая концентрация солей подавляет развитие корневой системы. В этом случае концентрацию солей уменьшают в 2-4 раза. Обычно для укоренения in vitro в качестве носителя используется агар. Однако агар не является инертным материалом, он содержит вещества, воздействующие на рост корней. Кроме того, твердая среда плохо аэрируется, из нее слабо удаляются продукты метаболизма.
Существуют 2 основных способа преодоления этих недостатков:
1) добавление активированного угля;
2) использование жидкой питательной среды в сочетании с верми-культурой или применением полиуретановой пены.
Возможно и укоренение в нестерильных условиях. Так, например, микропобеги голубики высокой можно укоренять в смеси верхового торфа с перлитом 1:3 после обработки нижней части микропобега ростовой пудрой. Укоренение производится в условиях повышенной влажности и притенения, т.е. этап 3 сочетается с этапом 4. Корни образуются в течение месяца.
Этап 4. Перенос в тепличные условия. Микрорастения, выращенные в культуре in vitro, имеют ряд анатомических и физиологических особенностей. Для них характерны более мелкие и тонкие листья, слабо развитая кутикула, нарушение работы устьиц, ксилемные ткани в регенерантах могут образовать закрытую систему, поскольку побеги появляются до образования корней. Кроме того, тип питания микрорастений миксо- или гетеротрофный, но не автотрофный. При пересадке в условия теплицы для адаптации микрорастения испытывают состояние стресса, что может привести к гибели части растений от недостатка влаги, повышенной температуры и других неблагоприятных факторов среды. В связи с этим главная задача этапа 4 - акклиматизация микрорастений в условиях in vivo. Важно также контролировать патогены, поскольку в условиях in vivo микрорастения могут инфицироваться бактериями, грибами, вирусами. В качестве дополнительной задачи этого этапа может быть укоренение, если корни слабо развивались на этапе 3. Для успешного решения названных проблем микрорастения пересаживаются в теплицы или комнаты с контролируемыми условиями. Главные требования успешной акклиматизации растений in vivo: высокая относительная влажность, уменьшенная интенсивность света, не слишком влажный субстрат и оптимальная температура. В теплице над пересаженными растениями устанавливаются укрытия из полиэтиленовой пленки для получения высокой влажности. Оптимальная влажность поддерживается с помощью туманообразующей установки, поскольку важен размер капель воды (чем меньше, тем лучше). Такие условия удерживают в течение 2-3 недель. Для предотвращения инфицированности микрорастений используют фунгициды и стерилизацию субстрата.
В качестве субстрата применяют верховой торф, а также его смеси с песком и/или перлитом. Хорошие результаты при укоренении микрорастений показал субстрат на основе ионообменных смол «Биона», разработанный в Институте физико-органической химии НАН Беларуси. Субстрат содержит необходимые элементы по прописи Мурасиге, Скуга. Важным его достоинством является возможность многократного использования. Микрорастения высаживают непосредственно в грунт или стеллажи теплицы, в ящики, горшки или пластиковые кассеты. Кассеты удобны в транспортировке и обеспечивают растения при пересадке стартовым запасом субстрата, сохраняя при этом корневую систему. Акклиматизация растений может начаться в культуре in vitro. Для этих целей открывают культуральные сосуды за несколько дней до высадки. Возможно также охлаждение дна сосудов.
Растения в естественных условиях вступают во взаимодействие с почвенной микрофлорой. На корнях многих растений образуется микориза, ряд растений взаимодействуют в ризосфере со свободноживущими микроорганизмами. Микроорганизмы почвы улучшают азотное и фосфорное питание растений, снабжают корни регуляторами роста (ауксины, цитокинины), подавляют развитие патогенов. Поскольку микрорастения in vitro не вступают в контакт с микроорганизмами, целесообразно создавать такие микробоценозы в ризосфере на этапе адаптации, для чего особенно эффективны пластиковые кассеты. Источниками микроорганизмов могут быть специально культивируемые штаммы, а также природные ассоциации. В БГСХА в течение ряда лет изучалась возможность использования микроорганизмов в клональном размножении. Установлено, что добавление ризофила и азоспириллы в субстрат в кассетах обеспечивает повышение массы одного растения на 28-48%. Азоспирилла способствует снижению заболеваемости растений на 16-23%.








