 2018-03-09
2018-03-09 1653
1653
Контролируется последовательностью нуклеотидов в ДНК-матрице.
РНК-полимераза доходит
до ЦГ-богатого участка,расплетание здесь идет очень медленно, что замедляет движение РНК-полимеразы.
Синтезированнаямолекула
РНК формируетстебель-петлю на участке богатом Г-Ц нуклеотидами, за которойрасположено несколько урацилов, что приводит к отсоединению моле-кулы РНК от матрицы ДНК

Rho - зависимый механизм терминации транскрипции
Rho-белок – гексамер.
Молекулярная масса
275 кДа.
Обладает хеликазной
актив 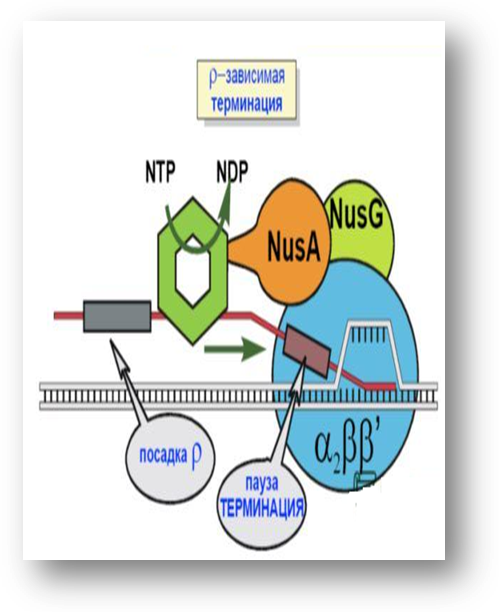

Транскрипция прокариот
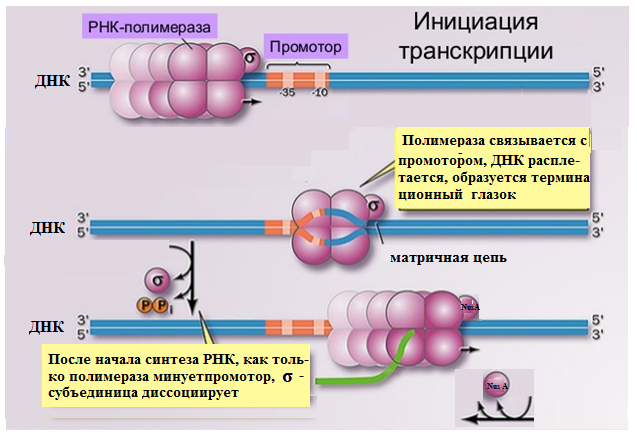
На первом этапе транскрипции у прокариотпроисходит связывание РНК-полимераз с молекулой ДНКипоискпромотора,на котором образуется инициаторный комплекс.
β′-субъединица обеспечивает свя-зываниехолофермента с молекулой ДНК до обнаружения промотора;-
σ-субъединица узнает промотортранс-криптона;
2α-, β- иβ′-субъединицы (α2 ββ′) служат для взаимодействия с промотором;
β-субъединица участвует в связывании рНМФвцепиРНК и удлинении молекулы РНК;
функция ω-субъединицы - восстанавление денатурированной РНК-полимеразы
Послеинициации синтеза РНКσ-субъединица отсоединяетсяот холофермента.
Дальнейший синтез РНК осуществляет корфермент, в который,на место σ-субъединицы встраивается фактор элонгации белок nus А, обеспечивающий эффективную элонгацию процесса.
ОбщийразмерРНК-полимеразы привзаимодействии с ДНКсоставляет 60 н.п.




В структуре РНК-полимеразы присутствует канавка длиной 55 Å (5,5 нм) и шириной 25 Å (2,5 нм). Именно в эту канавку помещается двойная спираль ДНК, имеющая ширину 20 Å (2 нм). На длине канавки укладывается 16 нуклеотидов.
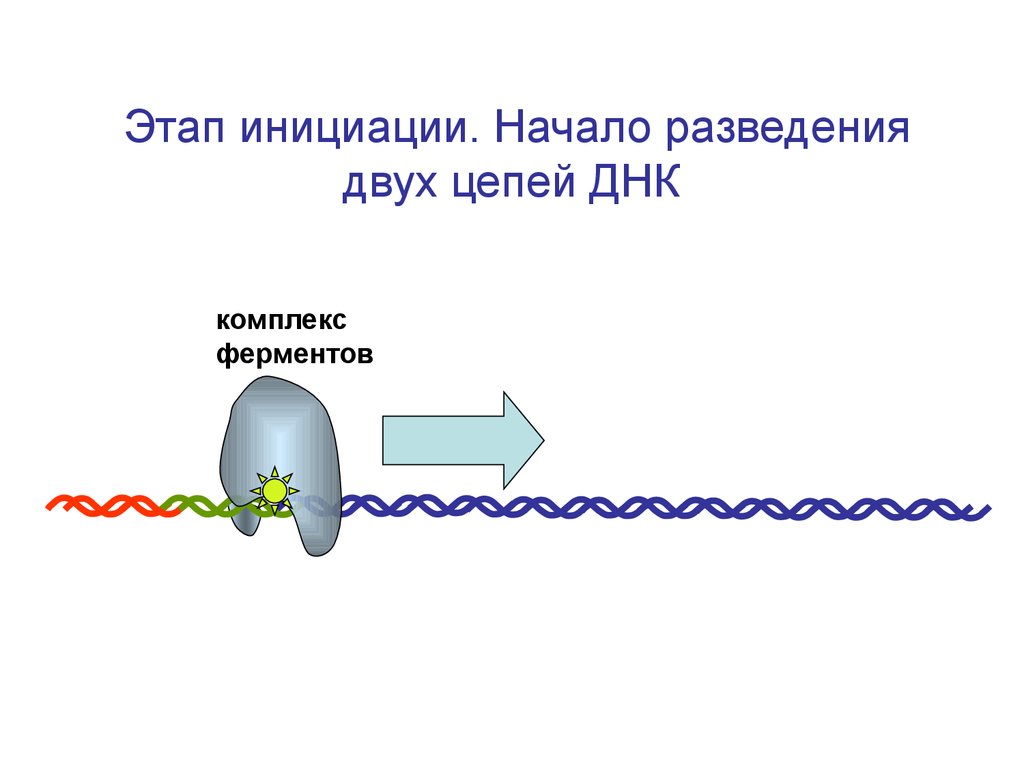
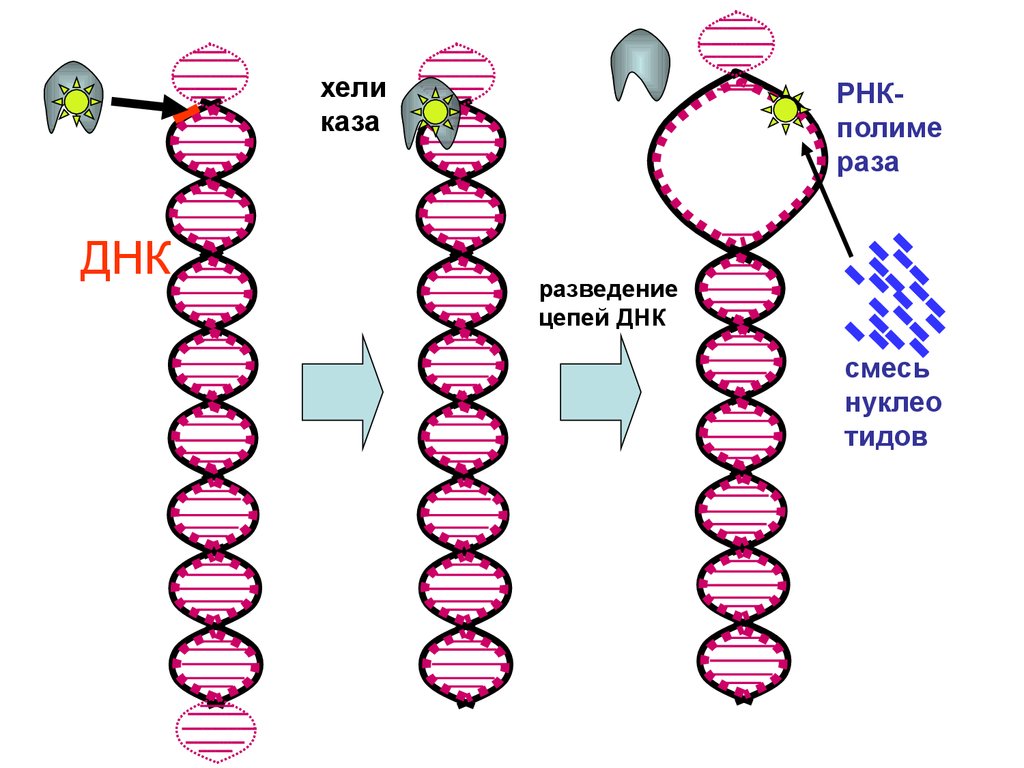




Элонгация транскрипции – это удлинение цепи за счет присоединения рибонуклеотидов. Онопроисходит по мере передвижения РНК-полиме-разы по ДНК. Соответственно, перемещается транскрипционный «гла-зок». После ухода «глазка» двуцепочная спираль ДНК сразу восстанав-ливается.
Синтез идет со скоростью примерно 30 нуклеотидов в секунду. В процессе элонгации возможны «ошибки» спаривания, то есть, встраивание «неправильных» нуклеотидов. Частота ошибок составляет 2∙104 нуклеотидов.

Терминация транскрипции – это окончание процесса синтеза РНК.
Окончание транскрипции связано с наличием специальных локусов ДНК, которые являются сигналами об окончании транск-рипции и называются стоп-сигналами.
Стоп-сигнал –это участок ДНК, богатый ГЦ –парами, за ним идет олиго-(А)- последовательность, содержащая 4-8 аденинапод-ряд.
Сила взаимодействия ГЦ пар очень сильна. Расплетание таких участков в ДНК прои-ходит трудней. Это замедляет ходРНК-полмеразы и является сигналом для прекращения транскрипции.
Транскрипция ГЦ-богатого участка приводит к образованию в РНК-транскрипте участка со вторичной структурой в виде «шпи-льки». «Шпилька» нарушает прочность связи РНК с молекулойДНКивытесняетРНК-полимеразу из термининационногоглаз-ка.Сразу, вслед за этим,РНК-транскрипт отделяется от молеку-лы ДНК и структура молекулыДНКвосстанавливается.



Транскрипция у эукариот
На 1 этапе транскрипции РНК-полимераза должна связаться с промотором. Наиболее детально этот этап изучен для РНК-полимеразы II, каталитирующей синтез мРНК.
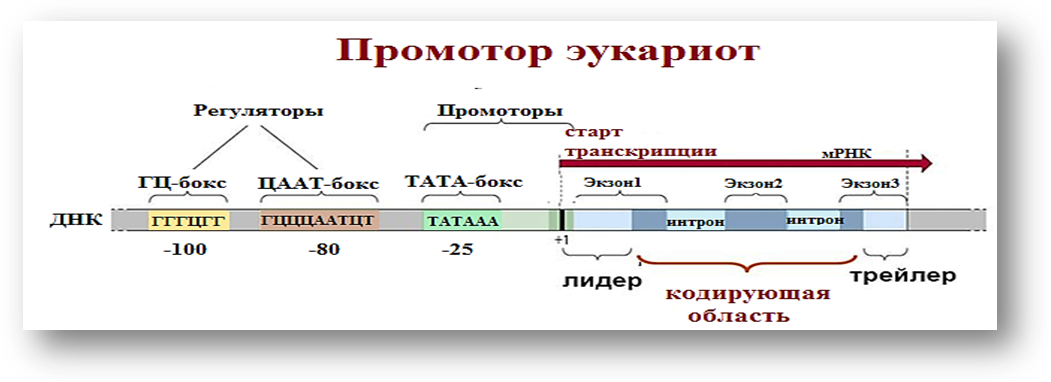
В общем плане, процесс транскрипции у эукари-от сходен с процессом транскрипцииу прокариот, но имеются инекоторые различия:
1.ДНКэукариот имеет нуклеосомное строение, поэтому неоходимо освободить её от гистонов в месте ее транскрипции.
2. В геноме эукариот нет оперонов. Регуляцию транскрипции осуществляют общие факторы тран-скрипции.
3. Медиатор – белковый комплекс, обеспечивающий взаимодействие регуляторных белков РНК-полимеразы
4. РНК-полимеразыуэукариот бывают 3 видов: РНК-полимераза I, II, III (далее pоl I, роl II, роl III);
5. Для эукариот характерно наличие энхансеров и сайленсероврегулирующих транскрипцию;
6. Процессингу подлежат все виды РНК:мРНК,тРНКирРНК;
7. 5 главных транскрипционных факторов (27 субъединиц)
Ни одна РНК-полимераза не способна самостоятельно связываться с промотором.
Для связиРНК-полимеразспромоторомимеются специальные белки – факторы транскрипции (ТF-факторы).
В зависимости от того, какую РНК-полимеразу связывают ТF-факторы, их называют ТFI, ТFII, ТFIII.
Латинская буква,следующаяза римской цифрой, обозначает, каким по счету был открыт этот фактор: ТFIIА, ТFIIВ, ТFIIД и т.д.
Различные факторы соединяются с молекулойДНК наразных сайтах
Например, ТFIIД соединяется с ТАТА-боксом, поэтому его называет ТАТА-фактором.На сегодняшний день известно шесть факторов транскрипции: ТFIIА, ТFIIВ, ТFIIД, ТFIIЕ, ТFIIF,ТFIIH.
На первом этапетранскипции происходит связывание РНК-по-лимераз с молекулой ДНКипоискпромотора,на котором образуетсяинициаторныйком-плекс.
Инициирующий комплекстранскрипции
Первым с промоторомтранскриптона связывается белок, узнающий ТАТА-бокс:ТАТА-связывающий белок ТВР.
С ТВР связываются еще 10 белков–ТВР-ассоциированныхфакторов или ТАF-белки (ТВР-associatedfactors).
ТВР-белок и ТАF-белки вместе составляют комплекс общих факторов транскрипции ТFIIД, II еще ряда активирующих транскрипцию комплексов: ТFIIВ, ТFIIF, ТFIIЕ и ТFIIН Каждый из этих комплексов выполняет свою функцию: ТFIIА (3 субъединицы) стабилизирует связь ТВР с промотором, ТFIIВ (1 субъединица)обеспечивает узнавание точки начала транскрипции; ТFIIF (2 субъединицы) стимулирует элонгацию, ТFIIН (7 субъединиц) обеспечиваетрасплетание спирали ДНК, то есть, обладает функцией хеликазы.Все эти комплексы функционируютвклеткахлюбого типаитакжеявляетсяобщимифакторами транскрипции.



 Образование инициирующего транскрипционного комплекса соп-ровождаетсяраскрытием двуцепочной молекулы ДНК, формируется транскрипционный глазок и начинается транскрипция.
Образование инициирующего транскрипционного комплекса соп-ровождаетсяраскрытием двуцепочной молекулы ДНК, формируется транскрипционный глазок и начинается транскрипция.
После образования молекулы РНКдлиной в 8-10 нуклеотидов,РНК-полимераза покидает промотор и начинается этап элонгации.
Последний момент инициации связан с вступлением в транскрипцию энхансеров и многочисленных факторов регулирующих скорость транскрипции. Эти фак-торы связываются с энхансером и транскрипционным комплексом.В результате образуется петля, сбли-жающаяэнхансер и промотор.
Скорость транскрипции = 100 нуклеотидов в секунду
Элогация цепи РНК у эукариот подобна таковой у прокариот:
· синтез РНКидет согласно принципу комплементар-ности;
· РНК –полимеразадвигает по матричной цепи ДНК в направлении 5′→3′.
· В молекуле РНК-полимеразы имеются центры связывания с молекулой ДНК и растущей цепью РНК;
· по мере движения РНК-полимеразы происходит локальные расплетание молекулы ДНК и восстановление ее структуры после ухода с этого участка фермента, то есть, транскрипционный глазок перемещается.
Точность спаривания нуклеотидов при транскрипции ниже точности репликации, так как, у РНК-полимераз отсутствует система коррекции ошибок;оналишена 3′→5′- и 5′→3′-экзонуклеазной активности. В результате частота ошибок составляет 1ошибку на 103-104 нуклеотидов.
Процесс элонгации у эукариотрегулируется группой белковых факторов элонгации:
· Р-ТЕFв - препятствует прекращению элонгации, вызванной действием ингибиторов;
· SII и ТFIIF контролируют активность РНК-полимеразы,как на стадии инициации, так и на стадии элонгации;
· SIII (элонгин) содержит 3 субъединицы, выполняющие функции активации и регуляции транскрипции, устранение задержки транскрипции. Мутация этого фактора приводит в возникновении многих видов рака у человека.
Терминация транскрипции у эукариот происходит также, как у прокариот в участках ДНК, богатых ГЦ-последовательностями. ОсвобождениеРНК-полимеразы и вновьсинтезированной РНКиз транскрипционного комплекса обеспечиваюттрифакторатерми-нации:
· для РНК-полимераза I-белки Rев-1 и ТТF-1,
· для РНК-полимеразы II- белок N-ТЕF,
· для РНК-полимеразы III-Lа-белок.
Все эти факторы связываются состоп-сигналамина молекуле ДНК и останавливают РНК-полимеразы. В этот момент, происхо-дятконформационныеизменения молекул РНК-полимераз и это приводит к ослаблению связей между участками транскрипционного комплекса и его распаду.
ОсвобождениеРНК и РНК-полимеразпроисходит при участии дополнительных факторов RТFR (polymeraseandtranscriptreleasefactors). Терминация завершается разрезанием РНК, после чего к её 3' концу фермент добавляет несколько аденинов (…АААА), от числа которых зависит стабильность данного транскрипта.
 Синтез РНК упро-,иэукариот имеет конвейерный характер; с одной транскрипционной единицы сопровождается созданием множества копий РНК.
Синтез РНК упро-,иэукариот имеет конвейерный характер; с одной транскрипционной единицы сопровождается созданием множества копий РНК.
После ухода РНК-полимеразы с про-мотора, с ним свя-зывается следую-щая молекула фер-мента и начинает транскрипцию но-вой молекулы РНК.
Среднее расстояние между несколькими ферментамиРНК-полимеразы составляет примерно 300-500 н.п. и зависит от «силы» промотора (во многом зависящей от транскрипционных факторов и активности РНК-полимеразы).
В результате транскрипции синтезируется молекулы – предшественникиактивныхРНК:пре-рРНК,пре-мРНКипре-тРНК, которые называются гя-РНК.
 Синтезированная в ядре гяРНК образует комплекс с глобуляр-ными ядерными белками – информоферами, частиц с массой30S.
Синтезированная в ядре гяРНК образует комплекс с глобуляр-ными ядерными белками – информоферами, частиц с массой30S.
Накаждый информофер накручивается отрезок гяРНК длиной в 600 нуклеотидов. Во время нахождения в ядре,гяРНК входит в состав информоферы, проходя процессинги превращаясь в зрелую мРНК.
Привыходеиз ядра в цитоплазму связь с информоферой нарушаетсяимРНК вступает в связь с белками цитоплазмы, выполняющимизащитную и транспортную функции, образуя информосомы – цитоплазматическиеРНП-частицы, открытые в 1964 году в лабораторииакадемикаА.С.Спиринапри участии академика АН КазССР М.А.Айтхожина.
Информосомыиграютопределённую роль в процессах трансляции РНК.
Пре-мРНК в несколько раз длиннее зрелых РНК, так каквключают транскриптыспейсеров ДНК и интроны, которые образуют «шпильки» Кроме того, длина пре-мРНК значительноварьирует у разных молекул от 2 тыс.п.н. до 20 тыс. п. н., поэтому её называют гетерогенной ядерной РНК (гя-мРНК). У нее отсутствует на 5′-конце «колпачок» (кэп), а на 3′-конце – поли (А) фрагмент.Пре-мРНК могут нести информацию об одном полипептиде или нескольких, расположенных на ДНК кластерами (гистоновые гены), и в этом случае пре-мРНК является полицистронной (в дальнейшем из нее образуется несколько зрелых м-РНК).
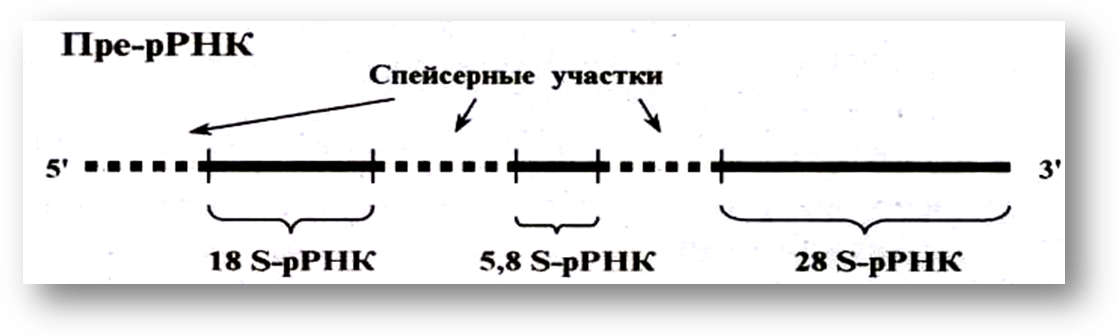
Пре-рРНКсодержит информацию о трех генах рРНК и, образуетзрелые р-РНК: 16S-,18S-;5S-,5,8S-;и 23S-,28SрРНК. Эти последовательности разделены спейсерами, но не содержат интронов
Пре-тРНКвсегдасодержат последовательность только одной зрелой тРНК. Она уже имеет вид «кленового листа», но не содержит ми-норных нуклеотидов, несформирован кодон акцепторной петли (ЦЦА) и антикодон не занимает своего правильного места.
Превращение пре-РНК в зрелые РНК осуществляется в ходе процессинга.
 Процессинг РНК - это процесс посттранскрип-ционных модификаций (изменений) предшественников РНК (пре-РНК).
Процессинг РНК - это процесс посттранскрип-ционных модификаций (изменений) предшественников РНК (пре-РНК).
У прокариот процессинг затрагивает только пре-рРНК и пре-тРНК. Пре-РНКпрокариот включают все виды рРНК и тРНК (рис.118), разделенные спейсерами.
Процессинг включает следующие преобразования:
· Кэпирование
· Полиаденилирование
· Сплайсинг
Процессинг у прокариот заключается в вырезании спейсеров ферментом РНКазой III, который является эндонуклеазой (ферментом, разрезающий нуклеиновые кислоты).
В результате образуются зрелые 16S-рРНК,23S-рРНК и 5S рРНК.
У пре-тРНК, после удаления спейсеров, процессинг продолжается при участии ферментов эндонуклеазы Р и эндонуклеазы Д.
Этиферментыудаляютлишние нукле-отиды в молекулепре-тРНКс 5'-, и 3'-концов; одновре-менно происходит модификация ряда азотистых оснований нуклеотидов в молекуле тРНК с образованием минорных нуклеотидов:уридин восстанавливается с образованием дигидроуридина; подвергается изомеризации с образова-нием псевдоуридина; метилируется с образованием метилуридина; аденин дезаминируется с образованием инозина и т.д.

