 2018-02-13
2018-02-13 2221
2221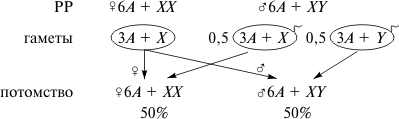
Такое наследование наблюдается при разных типах хромосомного механизма определения пола, и вероятность рождения потомков мужского и женского пола в норме одинакова. Однако баланс полов может быть нарушен, если в половых хромосомах возникают летальные мутации. Рассмотрим случай, когда рецессивная летальная мутация (l) возникла в одной из двух Х-хромосом самки дрозофилы (XBl), маркированной доминантной мутацией Bar (В) — полосковидные глаза. Рассмотрите схему скрещивания такой самки с нормальным самцом дикого типа (+), имеющим круглые глаза.
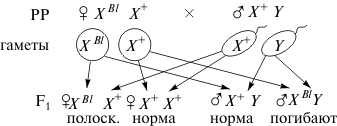
Как видно из схемы, появление рецессивной летальной мутации в одной из Х-хромосом самки приводит к гибели половины мужского потомства. Об этом судят по отсутствию самцов с полосковидными глазами, получившими от матери Х-хромосому с летальным геном (XBl).
Гены, определяющие признаки пола, имеются не только в половых хромосомах, но и в аутосомах. С другой стороны, признаки, которые наследуются сцепленно с полом, часто не имеют прямого отношения к полу. Существует особая категория признаков, которые проявляются только у одного пола. Это — ограниченные полом признаки. Определяющие их гены имеются у обоих полов и могут находиться как в половых хромосомах, так и аутосомах. Однако работают эти гены, т.е. проявляют свое действие на уровне фенотипа, только у одного пола. К числу таких признаков относятся, например, молочность и жирность молока у коров, яйценоскость и размер яиц у кур. Эти признаки, которыми обладают особи женского пола, могут целиком определяться генотипом отца. Такое явление широко используется в селекции животных при использовании отцовских особей-производителей для получения высококачественного потомства.
Гены, определяющие развитие вторичных половых признаков, имеются как у мужчин, так и у женщин, но их проявление контролируется гормонами.
Пол может оказывать влияние на характер проявления признака, т.е. на его доминантность или рецессивность. В этом случае признаки называют зависимыми от пола. Так, например, у овец ген, определяющий развитие рогов, является доминантным у самцов и рецессивным — у самок. В связи с этим гетерозиготные самки являются комолыми, а гетерозиготные самцы — рогатыми. У человека точно так же наследуется признак плешивости. Зависимые от пола признаки находятся под сильным влиянием половых гормонов, соотношение которых может либо усилить, либо ослабить экспрессию гена.
Итак, подведем итог, касающийся механизма определения пола. Пол, как любой другой признак организма, детерминирован генетически. В определении пола у большинства животных и растений основная роль принадлежит половым хромосомам. Расщепление по полу соответствует соотношению 1: 1, что обусловлено равновероятным образованием двух типов гамет (1/2 с Х и 1/2 с Y хр.) у гетерогаметного пола (XY). Гетерогаметным может быть как мужской, так и женский пол.
Определение пола — это начальный этап становления пола, за которым следует процесс его дифференциации, приводящий к развитию двух разных половых типов — женского и мужского. У животных половая дифференциация затрагивает всю организацию особи: строение органов размножения, внешнюю морфологию, обмен веществ, поведение, гормональный баланс, продолжительность жизни и пр. Половые различия которые обеспечивают комбинативную изменчивость внутри вида, а также его изоляцию, являются адаптивным механизмом.
Различают первичные и вторичные половые признаки. Первые непосредственно обеспечивают осуществление полового процесса. В частности, к ним относятся различия в строении внешних и внутренних половых органов женских и мужских особей. Развитие вторичных половых признаков является результатом нормального функционирования гонад (т.е. опосредовано первичными половыми признаками) и способствует половому размножению. Регулируется развитие вторичных половых признаков с помощью половых гормонов.
На процесс дифференциации пола оказывают влияние как генотипические факторы, так и внешняя среда.
Еще в начале ХХ в. было высказано предположение, что зигота является потенциально бисексуальной, но существуют механизмы, осуществляющие дифференциацию пола. Одним из таких механизмов является баланс половых хромосом и аутосом, при нарушении которого развитие пола отклоняется либо в сторону женского, либо в сторону мужского пола. Необходимость такого баланса впервые была установлена в опытах К. Бриджеса (лаборатория Т. Моргана), который обнаружил линию дрозофилы, дающую наряду с нормальными самцами и самками большой процент интерсексов. Интерсексы представляют собой смесь первичных и вторичных мужских и женских половых признаков, образуя все переходные типы: от сходных в основном с самцами до сходных с самками. Все они стерильны. В опыте Бриджеса они возникли в потомстве триплоидных самок, оплодотворенных нормальными диплоидными самцами, и содержали три набора аутосом и нормальное количество половых хромосом: 2Х+3А. Наряду с типичными интерсексами, в потомстве были представлены особи с гипертрофированными признаками женского пола — суперсамки (3Х+2А), и мужского пола — суперсамцы (XY+3X).
На основании этих результатов Бриджес пришел к выводу, что не само присутствие двух половых хромосом (XX или XY) определяет развитие пола, а баланс половых хромосом и гаплоидных наборов аутосом. Поскольку у дрозофилы Y-хромосома генетически инертна, то важно только количество Х-хромосом. Все особи с отношением 2Х: 2А = 1 являются самками, особи с отношением 1Х: 2А = 0,5 — самцами, типы с промежуточными между 1 и 0,5 отношениями являются интерсексами, а отношения больше 1 дают суперсамок, меньше 0,5 — суперсамцов.
Аномальное развитие пола при изменении числа наборов аутосом обусловлено нарушением баланса генов, которые участвуют в развитии пола. Поскольку гены проявляют свое действие в конкретных условиях, то на их функционирование оказывают влияние внешние факторы. Так, потомство триплоидных самок дрозофилы воспитывалось в условиях высокой и низкой температур. В обоих случаях развивались интерсексы, но при высокой температуре преимущественно с признаками самки, а при пониженной — с признаками самца. Таким образом, окончательное развитие пола является результатом сложных взаимодействий генов, локализованных как в половых хромосомах, так и в аутосомах, друг с другом и с факторами окружающей среды.
Изначальная бисексуальность зигот подтверждается фактами переопределения пола в процессе индивидуального развития. Классический пример — морской червь Bonellia viridis. Свободноплавающие личинки этого червя развиваются в самок. Если же личинка остается прикрепленной к материнской особи, из нее развивается самец. Будучи отделена от самки, такая личинка, начавшая развиваться в самца, изменяет направление дифференциации пола в женскую сторону и из нее развивается интерсекс. В хоботке самки имеются химические регуляторы, способные переопределять пол личинок.
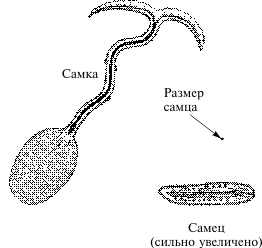
Половые различия у морского червя Bonellia viridis
Большой интерес представляет экспериментальное переопределение пола. Путем воздействия гормональными препаратами у ряда животных удается получить полное превращение пола вплоть до способности формировать половые клетки противоположного пола. Такое превращение известно у некоторых лягушек, рыб, птиц и других животных. Так, раннее удаление яичника у самок кур и голубей может изменить в мужскую сторону окраску оперения, поведение и даже вызвать развитие семенника. У крупного рогатого скота наблюдались случаи рождения разнополых двойнь, в которых бычок оказывался нормальным, а телка — стерильной, со многими чертами самцового типа. Такие двойни носят название “фримартинов”. Их появление обусловлено тем, что семенники мужского эмбриона рано начинают выделять мужской гормон, который попадает в кровь и оказывает влияние на близнеца.
Один из ярких примеров полного переопределения пола описан в 1953 г. японским ученым Т. Ямамото. Опыт проводился на белых и красных медаках (Oryzias latipes), у которых доминантный ген красной окраски находится в Y-хромосоме. При такой локализации гена при скрещивании самцы всегда будут красными, а самки — белыми. Фенотипических самцов кормили с добавлением в корм женского полового гормона. В результате оказалось, что все красные рыбки с генотипом самца являются самками с нормальными яичниками и женскими вторичными половыми признаками.
Переопределение пола может быть следствием мутаций отдельных генов, участвующих в дифференциации пола. Так, у дрозофилы в одной из аутосом обнаружен рецессивный ген tra, присутствие которого в гомозиготном состоянии обусловливает развитие женских зигот (XX) в фенотипических самцов, оказывающихся стерильными. Самцы XY, гомозиготные по этому гену, являются плодовитыми.
Аналогичные гены найдены у растений. Так, у кукурузы рецессивная мутация silkless в гомозиготном состоянии вызывает стерильность семяпочек, в связи с чем обоеполое растение функционирует как мужское. У сорго обнаружены два доминантных гена, комплементарное взаимодействие которых также вызывает женскую стерильность.
У наездника Habrobracon пол определяется по тому же типу, что и у пчел: диплоидные самки развиваются из оплодотворенных яиц, а гаплоидные самцы партеногенетически. Но иногда самцы могут развиваться из оплодотворенных яиц. Причина такой ситуации лежит в действии специфического гена, в гомозиготном состоянии определяющего развитие зиготы по мужскому типу.
Правильность хромосомной теории определения пола подтверждается существованием половых мозаиков, или гинандроморфов, совмещающих в себе части тела мужского и женского полов. Известны разные типы гинандроморфов: латеральные, переднезадние, мозаичные.

Билатеральный гинандроморф
Drosophila melanogaster
Латеральный гинандроморфизм описан у насекомых, у кур, у певчих птиц. В этом случае одна половина тела соответствует женскому типу, вторая — мужскому. При мозаичном гинандроморфизме большая часть тела имеет признаки одного пола, и лишь отдельные участки — признаки противоположного пола. Этот тип описан, в частности, у дрозофилы. Чаще всего причиной появления гинандроморфов является утрата одной из двух Х-хромосом в раннем дроблении зиготы с кариотипом самки (ХХ). Клетки с кариотипом Х0 обнаруживают признаки мужского пола. Чем раньше произойдет элиминация Х-хромосомы, тем больше участков мужского типа будет представлено в теле взрослой мухи. Обнаруживаются такие мозаики по проявлению рецессивных генов, которые в зиготе находились в гетерозиготном состоянии, но проявились фенотипически в клетках с кариотипом Х0.
Еще одной причиной гинандроморфизма может быть развитие зародыша из яйцеклетки с двумя ядрами (дизиготический гинандроморфизм). В этом случае мозаики могут быть соматическими, если оба ядра имеют один и тот же набор половых хромосом, но разный генотип (например, одно ядро Аа, а другое — аа), или половыми, если одно ядро ХХ, а другое ХY, или теми и другими одновременно. Подобный тип гинандроморфизма описан у шелковичного червя, бабочки, дрозофилы.
Известен также гинандроморфизм, причиной которого является полиспермия. Он обнаружен у дрозофилы. В яйцеклетке дрозофилы могут сформироваться два женских гаплоидных пронуклеуса, с одной Х-хромосомой каждый. При проникновении в яйцеклетку двух сперматозоидов один пронуклеус может оплодотвориться сперматозоидом с Х-хромосомой, а другой — сперматозоидом с Y-хромосомой. После первого дробления образуются два бластомера, один с кариотипом ХХ, другой — ХY, что в дальнейшем приведет к развитию гинандроморфа.








