 2020-04-20
2020-04-20 707
707
Функциональная неравнозначность клеток и связанная с ней репрессия и активация генов давно привлекали внимание генетиков, но до последнего времени реальный механизм контроля генной активности оставался неизвестным.
Первые попытки объяснить регуляторную активность генов были связаны с изучением гистонных белков. Еще супруги Стэдман в начале 40‑х годов нашего века высказали мысль, что именно гистоны могут выполнять роль контролеров активности генов. В своих дальнейших работах они получили первые четкие результаты о различиях в химической природе гистонных белков. Обобщая полученные данные, они писали: «Физиологические функции ядер являются, вероятно, следствием присутствия генов, которые они содержат. Они (гены – В.С.) должны поэтому быть идентичными во всех ядрах данного организма. Если, однако, предположить, что ядра содержат некоторый механизм для подавления активности отдельных генов или групп генов и что этот механизм специфичен для каждого типа клеток, эти трудности исчезнут. Продемонстрированные нами данные, что некоторые основные белки, имеющиеся в клеточных ядрах, являются, несомненно, клеточно‑специфичными, приводят к гипотезе, что одна из физиологических функций заключается в том, чтобы действовать в качестве репрессоров генов»[192]. Эта догадка получила некоторое подтверждение в более поздних работах Дж. Боннера, Ру‑Чи‑Хуанга, Тсьо и других.
Сейчас несколько коллективов исследователей интенсивно работает в этой области. Объем данных, свидетельствующих в пользу этой гипотезы, довольно велик, и можно полагать, что она получит экспериментальное подтверждение.
В то же время все большее количество фактов говорит за то, что регуляция генной активности гораздо более сложный процесс, нежели простое взаимодействие участков генов с молекулами гистонных белков. Об этом в первую очередь свидетельствуют эксперименты по регуляции генов у микроорганизмов (см. также главу 23).
В 1960–1962 гг. в лаборатории Р.Б. Хесина‑Лурье удалось выяснить, что гены фагов начинают считываться не одновременно. Было показано, в частности, что гены фага Т2 можно разделить на ранние, работа которых падает на первые минуты заражения бактериальной клетки, и поздние, начинающие синтезировать иРНК после завершения работы ранних генов.
Четкая координированность действия генов и их своеобразная иерархия была доказана Ф. Жакобом и Ж. Моно (1961). Они показали, что гены бактерий можно условно разделить на два различных типа – структурные гены, дающие информацию о синтезе определенных белков (ферментов), и регуляторные гены, следящие за включением и выключением отдельных генов или их блоков в зависимости от метаболических потребностёй клетки. По представлениям Жакоба и Моно, регуляторные гены должны детерминировать особые молекулы репрессоров, которые, соединяясь с другими генами регуляторной системы – генами‑операторами, управляют работой последних. Тем самым Жакоб и Моно разделили гены регуляторной системы в свою очередь на два типа – гены‑регуляторы и гены‑операторы[193]. В экспериментах с кишечной палочкой они смогли дать принципиальное доказательство существования этих типов. По их данным, ген‑регулятор находится на некотором отдалении от структурных генов, управляемых им, а оператор непосредственно к ним примыкает. Авторы ввели в генетику новое понятие, определив блок структурных генов и управляющий ими оператор, как единую функциональную единицу – оперон.
В последние годы были получены данные о наличии еще одной управляющей ячейки генной активности – промоторе. Оказалось, что по соседству с операторным участком, к которому присоединяется продукт – белковое вещество репрессор, синтезированный на гене‑регуляторе, имеется другой участок, который также следует отнести к членам регуляторной системы генной активности. К этому участку присоединяется молекула фермента РНК‑полимеразы. В этом промоторном участке должно произойти взаимное узнавание уникальной последовательности нуклеотидов в ДНК и специфической конфигурации белка РНК‑полимеразы. От эффективности узнавания будет зависеть осуществление процесса считывания генетической информации с данной последовательности генов оперона, примыкающего к промотору.
Таким образом, схема взаимодействия гена‑регулятора, оператора, промотора, ферментов и рибосом, участвующих в синтезе специфических белков, может быть представлена следующим образом (см. рисунок): на гене‑регуляторе синтезируется вещество белковой природы – репрессор. Последний может воздействовать на операторный участок – выключать (или в некоторых случаях включать) его. Репрессор, кроме того, может взаимодействовать с метаболитами, синтезируемыми под контролем специфических генов. Если метаболиты накопились в достаточном количестве, они, взаимодействуя с репрессором, изменяют его конфигурацию, и он отсоединяется от операторного участка. В том случае, когда оператор находится в состоянии, при котором он и находящиеся под его контролем структурные гены могут функционировать, к промоторному участку присоединяется молекула РНК‑полимеразы, начинающая считывать информацию с данного оперона. Информация считывается в виде последовательности оснований в молекуле иРНК. К концу этой иРНК присоединяется рибосома и, продвигаясь вперед, считывает информацию с иРНК, в результате чего синтезируется полипептидная цепь. Вслед за первой рибосомой присоединяется вторая, за ней третья и т. д. Таким образом осуществляется взаимно скоординированное функционирование частей белок‑синтезирующего аппарата.
Такова картина синтеза белка в бактериальных клетках. В клетках высших организмов она оказалась несколько иной: молекулы иРНК после окончания их синтеза отделяются от матрицы ДНК и скапливаются в цитоплазме. Предполагают, что они переходят в цитоплазму соединенными со специальными белками (возможно, близкими к гистонным белкам).
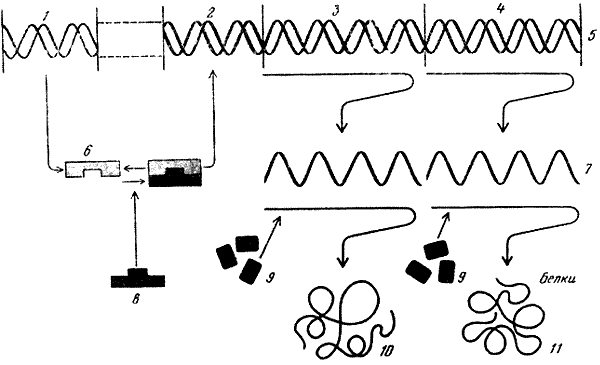
Схема генетической регуляции белкового синтеза, по Жакобу и Моно (1961).
1 – регуляторный ген; 2 – оператор; 3 – структурный ген А; 4 – структурный ген В; 5 – ДНК; 6 – репрессор; 7 – РНК; 8 – регуляторный метаболит; 9 – аминокислоты; 10 – фермент А; 11 – фермент В.

Схема, поясняющая взаимодействие регуляторных и структурных генов при синтезе белка (по Жакобу и Моно, 1961).
ГР – ген‑регулятор; П – промоторный участок; О – операторный участок; СГ1, СГ2 и СГ3 – первый, второй и третий структурные гены, входящие в данный оперон.








