 2020-05-12
2020-05-12 244
244
Изучение особенностей поведения рыб в токсических растворах, содержащих различные компоненты промышленных сточных вод, представляет интерес в первую очередь в связи с необходимостью всестороннего описания картины действия токсического вещества и внешне обнаруживаемых симптомов отравления рыб. Детальная характеристика симптомов отравления и их полный перечень могут быть использованы в качестве индикаторов для определенных групп ядов, что позволит устанавливать причину гибели рыб в естественных водоемах.
Знание симптоматологии отравления необходимо также для унификации показателей (при оценке токсического действия того или иного вещества), используемых в различных лабораториях. Между тем даже при изучении токсичности какого-либо одного вещества в разных лабораториях в качестве показателей его токсичности используют различные симптомы отравления без учета их специфичности для данной интоксикации, времени появления и продолжительности действия. Все это затрудняет понимание и сопоставление литературных данных, а подчас и полностью обесценивает их.
Сказанное в первую очередь относится к работам, в которых рассматривается токсичность органических соединений. Известно, что вещества этой группы ядов оказывают на организм более разнообразное влияние, вследствие чего внешние признаки отравлений этими веществами значительно разнообразнее в сравнении с симптоматологией токсического процесса, обусловленного ядами неорганической природы (тяжелые металлы, кислоты. Щелочи и др.). Это весьма затрудняет правильный выбор основных показателей токсического процесса, на основе которых можно судить о степени вредности исследуемого вещества.
В качестве примера рассмотрим общую картину и внешний симптомокомплекс фенбльного отравления рыб, поскольку гомологи фенольного ряда представляют собой группу ядов, широко распространенных во внутренних водоемах различных стран. Кроме того, peaкция рыб на яды фенольного ряда по качественному составу симптомокомплекса имеет много общего с реакцией рыб на многие другие компоненты промышленных сточных вод органического происхождения.
Общая характеристика фенольного токсического процесса и поведения рыб при отравлении различными концентрациями яда описывалась многими авторами [692, 792, 542].
Довольно подробно внешние симптомы фенольного и крезольного отравления были рассмотрены К. Масон-Джонсоном [692] у окуней и форели. Автор отмечает, что при высоких концентрациях яда у рыб наблюдались быстрая потеря рефлекса равновесия, «взбешенное плавание» и опрокидывание на бок. Дыхание, глубокое и учащенное вначале, по мере развития отравления становилось нерегулярным и слабым. В опытах М. Эллиса [542] на золотых рыбках в качестве характерного признака фенольного отравления был отмечен паралич нейромускулярного аппарата [861, 862, 630, 469, 40, 413, 370].
Эти наблюдения в основных чертах были подтверждены. Исследования К. Вурмана и X. Вокера [861, 862], проведенные на форели, пескарях, окунях, головлях, плотве к усачах, а также Д. Джонса [630] — на головлях, выявили следующие особенности поведения испытуемых рыб в слабых растворах фенолов: пониженную интенсивность двигательной активности, кратковременную потерю рефлекса равновесия с последующим восстановлением. Важной особенностью этих работ явилось значительное расширение видового состава испытуемых рыб.
Из работы В. Букштейга и др. [469] следует, что поведение рыб (окунь, гольяны) в растворах гидрохинона, пирогаллола и пирокатехина весьма сходно с их поведением при фенольном и крезольном отравлении. Авторы отмечали у рыб беспорядочные броски и сильнейшую дыхательную деятельность. Впоследствии рыбы заваливались на бок и оставались в таком положении до наступления смерти. Обращает на себя внимание особенность реакции угрей на токсическое действие фенолов. Они обычно остаются на месте при развитии интоксикации, причем удается отметить только «вздрагивание» без обычно развивающейся у других видов рыб бурной двигательной активности.
Привлекают внимание работы Е. А. Веселова [40] и М. Ф. Вернидуб [37а], в которых помимо описания общего поведения рыб в токсических растворах фенолов представлена развернутая картина фенольной интоксикации. Е. А. Веселое считает, что симптомы отравления карасей развиваются в определенной последовательности и можно выделить семь этапов от начала отравления до смерти рыбы. Характерными признаками отравления рыб фенолами автор считает паралич мышц туловища. Это подтверждается более ранними наблюдениями Н. А. Мосевича, А. Г. Гусева и М. Г. Драгулина [240], согласно которым одним Из важнейших признаков фенольной интоксикации у рыб служит кровоизлияние в мышцах туловища.
Литературные данные позволяют в общих чертах составить представление о характере реакции рыб в ответ на фенольное раздражение. Дальнейшее развитие работ в этом плане ставило перед необходимостью дать точную характеристику картины интоксикации с детальным учетом характерных признаков и их сменььво времени. Кроме того, для сравнения результатов отдельных опытов по выявлению токсичности различных фенолов-желательно было выделить наиболее существенные реакции, характеризующие качественно различные стадии отравления рыб.
С этой целью был проведен анализ общей картины и особенностей течения фенольной интоксикации 22 различных видов рыб (карась, лещ, плотва, густера, сипец, жерех, чехонь, радужная и/ ручьевая форель, окунь, судак, ерш, щука, осетр, стерлядь, сом, красноперка, вобла, тарань, севрюга, белоглазка и налим) при весьма широкой амплитуде концентраций токсиканта (от 5 до 800 мг/л). Выявлен однотипный (за редким исключением) характер реакции различных видов рыб па патогенное действие фенолов (фенол, орто-, мета- и паракрезолы, пирогалол). Детальная характеристика поведения рыб в токсическом растворе и внешние симптомы отравления могут быть представлены в следующем виде. Через 3-10 мин после погружения рыб в токсический раствор у них в зависимости от концентрации яда отмечается чрезвычайно бурная и беспорядочная двигательная активность со стремительными бросками из толщи раствора на поверхность, кратковременным заваливанием на спину и па бок с последующим выходом из бокового положения. Через 30-90 мин кратковременное заваливание на бок сменяется стойким нарушением рефлекса равновесия. В боковом положении рыбы находятся до наступления смерти. В начале отравления, когда рыбы находятся в относительно неподвижном Г боковом положении, у них отмечаются интенсивные клонические судороги с частотой 12-24 судорожных подергиваний в минуту, вслед за чем рыбы выходят из неподвижного состояния и начинают импульсивное толчкообразное перемещение в боковом положении.
Характер двигательных реакций бывает двояким: перемещение на поверхности раствора по кругу или неориентированные перемещения в толще раствора. Этот период развития фенольного токсического процесса (потеря рефлекса равновесия, опрокидывание на бок и импульсивное перемещение в боковом положении) является самым длительным и одним из наиболее характерных признаков-фенольного отравления.
Через некоторое время двигательная активность рыб в боковом положении постепенно затухает и полностью прекращается. Рыбы становятся неподвижными, движения всех плавников, кроме грудных, парализуются, отмечаются редкие клояические судороги, и рыбы погибают. Отметим, что даже в этот период полной обездвиженности испытуемых рыб их реакция на механические раздражители сохраняется, хотя и в значительно ослабленной форме.
Касаясь других признаков и реакций, составляющих внешний симптомокомплекс фенольного отравления, можно указать, на характерное дугообразное изгибание туловища рыбы вследствие одностороннего тетанического сокращения мышц хвостовой части, являющейся движителем рыб, а также на резкую пигментацию (потемнение) спины и головы с одновременным побледнением-брюшной и хвостовой частей туловища. Нередко, особенно при длительном воздействии различных концентраций яда, отмечались обширные геморрагические явления в области грудных и брюшных плавников.
Существенные изменения на всем протяжении токсического процесса претерпевает дыхательная функция рыб. В период общего двигательного возбуждения дыхание резко учащается (до 128-144 против исходных 48-60), а затем частота дыхательных движений оперкулярных крышек постепенно уменьшается, возвращаясь к исходной и ниже. Резко нарушается структура дыхательных движений, дыхание становится аритмичным и нерегулярным.
Таким образом, внешние проявления действия токсического раствора на организм рыб довольно разнообразны, но совокупность реакций стереотипно воспроизводится при действии различных концентраций яда. В общефизиологическом плане можно выделить две стадии: стадию возбуждения, сопровождающуюся первоначальной бурной двигательной активностью, потерей рефлекса равновесия и импульсным перемещением рыб в боковом положении, и стадию угнетения, включающую потерю двигательной активности, угнетение дыхания, потерю ориентировочной реакции и резкое ослабление чувствительности рыб к механическим раздражителям.
В практическом отношении необходимо учитывать основные наиболее важные реакции внешнего симптомокомплекса отравления рыбы. Ближайший анализ показывает, что постоянство внешнего симптомокомплекса фенольного отравления проявляется в обязательной смене трех следующих друг за другом фаз: резкая двигательная возбудимость с кратковременным заваливанием на бок; потеря рефлекса равновесия, опрокидывание на бок, импульсивное перемещение в боковом положении; боковое положение, сопровождаемое конвульсивными судорогами, завершающееся полной потерей двигательной активности, расстройством дыхания и смертью. Протяженность отдельных фаз во времени при разных концентрациях яда и соотношение отдельных фаз весьма вариабельны, однако 3-фазовая стандартная форма реакции сохранялась в опытах с большинством видов рыб. Вместе с тем при прочих равных условиях длительность и степень выраженности отдельных фаз определяются видовыми особенностями испытуемых рыб. Как правило, у мирных рыб (карась, лещ, плотва, синец) различные фазы фенольной интоксикации более продолжительны, чем у хищных (щука, налим) и полухищных (ерш, окунь, форель). Несколько иные, чем у остальных исследованных видов рыб, реакции внешнего симптомокомплекса у налима и сома, специфика которых в токсических растворах состоит в конвульсивных подергиваниях мышц туловища при неподвижном состоянии рыбы. Только изредка у этих видов рыб наблюется заметное повышение двигательной активности, лровождаемое резкими стремительными бросками. Стереотипное воспроизведение 3-фазовой фенольной-токсикации при отравлении различных видов рыб едставляет определенный интерес не только для унификации показателей токсического действия ядов фенольного ряда при изучении их токсичности в различных лабораториях, но и для интерпретации результатов опытов с другими ядами органической природы, в частности инсектицидами. Известно [571], что при отравлении рыб, например эндрином или токсафеном, вначале отмечается повышенная двигательная активность, затем — потеря равновесия, опрокидывание на бок, расстройство дыхания и смерть. Специфические особенности действия разных концентраний одного яда и одной концентрации различных ядов находят свое выражение в количественном варьировании неспецифической формы реагирования, которой растеризуется однотипная 3-фазовая реакция разных видов рыб. В этом отношении реакция рыб сходна с 3-фазовым изменением уровня защитно-адаптационного потенциала у теплокровных под влиянием разнообразныx чрезвычайных раздражителей (длительное голодание, воздействие низких температур, различных ядов, мышечное перенапряжение и т. д.), изменяющих постоянство внутренней среды организма.
В связи с этим рассмотрим основные положения учения Г. Селье [302] о стрессе как теории, намечающей перспективные пути обобщений и анализа характера реакций различных групп животных на самые разнообразные внешние влияния, нарушающие постоянство внутренней среды (гомсостаз). Термин «стресс» был перенесен Г. Селье в биологию из физики, где он характеризует напряжение как производное силы и сопротивления. С точки зрения биолога «стресс» означает состояние организма, обусловленное взаимодействием реакции защиты и повреждения. «Стресс — это состояние, выражающееся специфическим синдромом, — пишет Селье, — который состоит из всех вызванных неспецифических изменений в биологической системе» (с. 54).
Сами агенты (факторы), вызывающие состояние напряжения (стресса) получили название стрессоров, или стрессирующих воздействий. В основе теории стресса лежит экспериментально обоснованное представление об общем адаптационном синдроме (ОАС). Не останавливаясь на всех симптомах, характеризующих ОАС, число которых постоянно растет и, видимо, еще далеко от своего завершения, отметим, что в основе развития ОАС Г. Сельс четко различает три фазы: реакция тревоги (алармная реакция); фаза резистентности и фаза истощения (рис. 1), которые мы имели возможность наблюдать и в наших опытах на рыбах. Кратко характеризуя каждую фазу, необходимо отметить следующие интересующие нас моменты:
1) реакция тревоги (alarm reaction) представляет собой комплекс реакций, развивающихся в неадаптированном организме под влиянием стрессора и сообщающих ему состояние резистентности;
2) фаза резистентности характеризуется повышенной устойчивостью организма к действию стрессоров и приобретению организмом состояния адаптации;
3) фаза истощения сменяет фазу резистентности при длительной экспозиции организма к действию стрессоров.
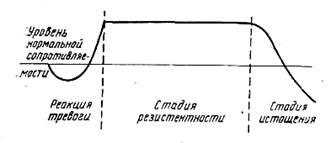
Рис. 1. Схема фраз общего адаптационного синдрома [302]
По существу, это три фазы общего адаптационного синдрома. Г. Селье характеризует динамику резистентности под влиянием стрессирующих факторов. Предпринятый нами экспериментальный анализ на модели фенольного отравления карасей [156] позволил вскрыть параллелизм между динамикой уровня резистентности и развитием фенолыюй интоксикации. Так, например, при действии на рыб'относительно низкой летальной концентрации фенола (25 мг/л) схема 3-фазового развития токсического процесса реализуется полностью и рыбы погибают в фазе истощения на 5—7-е сутки непрерывного действия токсиканта. Многократное увеличение силы токсического раздражителя (до 800 мг/л) приводит к быстротечной гибели всех подопытных рыб в фазе реакции тревоги, т. е. фаза адаптации не успевает проявиться даже у наиболее резистентных рыб, как это имеет место в опытах с летальными концентрациями фенола в диапазоне от 50 до 200 мг/л.
Обнаруженное нами 3-фазовое течение токсического процесса у рыб на уровне целостного организма было подтверждено впоследствии как в опытах с фенолом [395, 396, 220, 216], так и с другими ядами органического ряда — пестицидами [216, 317, 135] и нефтепродуктами [117]. Установлено, что фазовое течение токсического процесса основано на нарушении нормального хода физиологических реакций и биохимических процессов. Результирующей этих изменений является смерть подопытных рыб.
В серии опытов, выполненных на карпе, в качестве стрессовых факторов были выбраны фенол, сернокислый никель и азотнокислый хром, а также высокие и низкие температуры и дефицит кислорода [395, 396]. Показателями реакции организма служили уровень аптиокислительной активности структурных липидов, их качественные и количественные изменения как субстрата неферментативного (свободнораднкалыюго) окисления и, наконец, количественные изменения ряда веществ (перекисей, хинонов), сопровождающих это окисление. Выявлен сходный тип изменения антиокислитслыюй активности и уровня содержания свободных радикалов в тканях рыб (печень, жабры, селезенка) на воздействие различных по своей природе, чрезвычайных по силе раздражителей.
Однотипность изменений исследуемых параметров позволила авторам рассматривать их в качестве нсспецифических механизмов авторегуляции, представляющей собой защитную реакцию рыб. Она осуществляется путем изменения концентрации тканевых окислителей, которые в свою очередь регулируют концентрацию свободных радикалов, а следовательно, и продуктов свободнорадикального окисления. В применении к рыбам эта схема авторегуляции является оригинальной и согласуется с общепринятым мнением о месте и роли антиокислителей и свободных радикалов в авторегуляции теплокровных организмов [87, 119].
В общей для всех исследованных тканей динамике изменения антиокислительной активности липидов В. И. Чернышев и М. М. Телитченко [396] выделяют три характерных периода. Первый и второй периоды отражают авторегуляторные механизмы, обеспечивающие физиологически допустимые изменения в соотношении уровня свободнорадикальных реакций и ингибиторов этих реакций (антиокислителей). Третий период — повышенной антиокислителыюй активности — имеет место только в опытах с летальным исходом. Это реакция клеточных и тканевых систем авторегуляции на вступление вторичного фактора — эндогенных токсинов (продукты свободнорадикального окисления), который и определияет патофизиологическую картину отравления.
Первая и третья фазы динамики антиокислительной активности липидов сопровождаются накоплением е исследованных тканях таких биологически активных веществ, как перекиси и лизосоединения (лизолецитины, лизокефалины). Интересно, что усиление активности-свободнорадикальных процессов и увеличение биологически активных (токсичных) продуктов метаболизма липидов совпадают во времени со снижением стойкости эритроцитов [395, 396, 317].
Совокупность полученных к настоящему времени материалов, характеризующих физико-химические механизмы патологических процессов, позволяет приблизиться к пониманию реакций рыб на стресс-факторы на молекулярном уровне. При этом важно подчеркнуть, что ' неспецифическая 3-фазовая ответная реакция рыб на этом уровне принципиально'сходна с реакцией теплокровных животных на различные по природе етрессор-ные воздействия.
Рассмотрим данные, характеризующие реакцию рыб на системном уровне. Как известно, -целостность организма, нормальный ход его жизнедеятельности и сохранение постоянства внутренней среды (гомеостаза) обеспечиваются согласованной деятельностью двух регуля-торных систем: нервной и эндокринной. Особая роль в реализации защитных и приспособительных реакций, т. е. в сохранении гомеостаза, принадлежит гипоталамо-гипофизарной нейросекреторной системе (ГГНС.) [271]. Чрезвычайные по силе раздражители, в том числе и токсиканты, приводят к активации деятельности ГТНС в усиленному выведению нейрогормонов в общую циркуляцию теплокровных животных.
Исследованиями последнего десятилетия (220, 215, 216, 96] выявлены изменения структур нейроэндокринной системы, характерные для типичной стрессорной реакции, развивающиеся у рыб под влиянием различных токсикантов. Детальное изучение этого вопроса выполнено в лаборатории А. Л. Поленова, В. Е. Матей [2161 на модели фенольной интоксикации гуппи (Lebistes reticulatus). Ответная реакция ГТНС имеет четко выраженный 3-фазовый характер, соответствующий схеме общего адаптационного синдрома Селье.
Весьма показательна в этом отношении динамика изменений состояния ГГНС гуппи, выявленных в опытах с раствором фенола концентрацией 12,5 мг/л. На первой фазе развития токсического процесса происходит быстрое снижение содержания нейросекреторного вещества (НСВ) в ГГНС, обусловленное, по-видимому, активным выведением нейрогормонов в кровяное русло (реакция тревоги). Вторая фаза реакции гуппи на токсикант проявляется депонированием НСВ в ГГНС, т. е. накоплением нейросекрета во всех отделах этой системы (фаза адаптации). На заключительной, третьей фазе развития токсического процесса (фаза истощения) содержание НСВ в системе вновь резко снижается и к концу 6-месячных опытов отмечены признаки истощения ГГНС.
В. Е. Матей [216] обозначила соответствующие фазы морфофункционального состояния ГГНС в процессе развития фенольного отравления рыб фазами функциональной активности, депонирования и функционального напряжения. Выраженность каждой из трех фаз реакции ГГНС на токсическое воздействие и их продолжительность определяются концентрацией токсического вещества, т. е. аналогично тому, что отмечено нами в опытах на целостном организме [156]. Например, реакция ГГНС гуппи на остротоксичные концентрации (16,7 и 25 мг/л) носила однотипный характер, проявившийся в последовательной смене фаз депонирования секрета, функционального напряжения и истощения, завершающейся гибелью рыб.
Обращает внимание отсутствие фазы функциональной активности, т. е. первоначального резкого снижения НСВ в ГГНС, соответствующего реакции тревоги общего адаптационного синдрома Селье, которая имела место при воздействии па рыб более низкой концентрации фенола (12,5 мг/л). По мнению автора, эти различия могут быть объяснены двояко: либо при воздействии остротоксичных концентраций эта фаза вообще отсутствует и сразу развивается торможение ГГНС, либо она развивается более скоротечно и оказалась необнаруженной при заданной (10-дневной) частоте проведения анализов. Мы склонны разделить второе предположение.
Наконец, в опытах с остролетальной концентрацией фенола (40 мг/л), вызывающей гибель подопытных рыб в течение 96 ч, в ГГНС гуппи развивались изменения, свойственные типичной стрессорной реакции: первоначальное торможение процесса выведения и транспорта нейросекреторного вещества из перикарионов нейросек-реторных клеток («депонирование») с последующей нарастающей потерей нейросекрета из ГГНС, завершающейся полным опустошением ГГНС и гибелью рыб. Однотипные изменения морфофункциопалыюго состояния ГГНС отмечены и при воздействии других стрессорных раздражителей, в том числе летальных концентраций хлористого натрия и полихлорпинена. Эти факты однозначно свидетельствуют о неспецифичности регистрируемых реакций со стороны ГГНС на стрессорные воздействия различной природы. Наконец, и это очень важно, обнаруженные в опытах на рыбах изменения функционального состояния ГГНС в ответ на стрессорные воздействия аналогичны тем, которые имеют место у теплокровных животных и человека [271], и, следовательно, рыбы отвечают на стрессорные воздействия таким же путем, что и млекопитающие.
Таким образом, полученные в последнее десятилетие экспериментальные материалы подтверждают правомерность применения теории стресса в ихтиотоксикологии, поскольку реакция рыб на различные токсиканты сходна с общим адаптационным синдромом Селье не только феноменологически, но и по механизмам реализации. Фазовые изменения функционального состояния гипоталамо-гипофизарной нейросекреторной системы рыб являются важнейшим неспецифическим звеном ответной реакции на стрессорные воздействия. В условиях стресса, вызванного различными группами токсикантов, происходят активация гипофиза и освобождение адренокор-тикотропного гормона (АКТГ), стимулирующего синтез и выделение кортикостероидов из интеррепаловой железы, представляющей собой гомолог надпочечников млекопитающих. Получены прямые экспериментальные данные, свидетельствующие об изменении секреции и уровня содержания в кропи кортикостероидов, в частности кортизола, при разных токсических воздействиях на рыб [568, 512, 525, 243]. Иными словами, и эндокринная система рыб реагирует на стрессорные воздействия токсической природы так же, как эндокринная система теплокровных животных.
Особое значение теории стресса для ихтиотоксикологии определяется тем, что она дает основание рассматривать любой токсический процесс как совокупность реакций защиты и повреждения, раскрывает фазность изменений резистентности и указывает на факторы, изменяющие защитпо-адаптационный процесс, представляющий собой физиологическое содержание феномена резистентности. Наряду с этим анализ экспериментальных данных в свете теории стресса позволяет увязать исследования по токсикологии рыб с общим направлением исследований по приспособительным реакциям организма в ответ на самые разнообразные чрезвычайные раздражители, вызывающие нарушения гомеостаза.
Показательна в этом отношении работа X. Денцера [519]. Автор, оценивая токсическое действие ряда химических веществ по потреблению кислорода, частоте и глубине дыхания, приходит к выводу, что эти показатели изменяются под влиянием разнообразных ядов так же, как изменяется частота дыхания радужной форели под влиянием высоких температур (рис. 2). X. Дениер выделяет в сложной и многокомпонентной реакции рыб на повреждающие воздействия несколько фаз (см. рис. 2). Первые две из них: бескомпенсационная фаза I и фаза полной компенсации II — представляют собой нормальные биологические реакции, а две последующие: фаза неполной компенсации III и критическая фаза IV — отражают патологические сдвиги, наступившие в организме. В соответствии с этим X. Депцер предлагает различать порог нормальной реакции 1, порог повреждения 2, критический порог 3 и летальный порог 4. При определенной концентрации яда наступает такой момент, когда выходит из строя регуляторный механизм, компенсирующий повреждения организма, и наступает гибель рыбы.
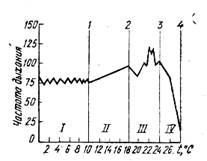
Рис. 2. Кривая изменения частоты дыхания радужной форели при повышении температуры воды.
К сожалению, такого рода анализ опытных данных, направленный на выявление общих и специфических черт в действии на организм рыб различных повреждающих агентов и прежде всего токсических компонентов промышленных сточных вод, весьма редок. Гораздо чаще авторы ограничивались описанием общего впечатления от поведения рыб в токсических растворах и некоторых типичных признаков наступившего отравления, которые удается выявить в результате внешнего осмотра погибших рыб. Известно, что поведение рыб в растворах, содержащих различные химические вещества, имеет существенные особенности. Например, растворы аммиака, фенолов и некоторых других ядов органического происхождения оказывают возбуждающее действие на рыб, сопровождающееся характерной бурной двигательной активностью, в то время как соли тяжелых металлов и прежде всего соли меди действуют на рыб угнетающе.
Следует отметить, что даже при воздействии одного и того же яда внешние признаки отравления погибших рыб могут быть неодинаковы. Согласно нашим данным внешние симптомы отравления фенолом мирных рыб, с одной стороны, и хищных и полухищных, с другой, различны. В момент, гибели карповых у них обычно слабо открыт рот и прикрыты жаберные крышки. Все плавники расправлены, а мышцы туловища находятся в состоянии тонического напряжения, которое значительно превосходит окоченение, наблюдающееся также при отравлении аммиаком. Большинство видов рыб после гибели обесцвечивается в течение ближайших 0,5-2 ч, что совпадает с более ранними наблюдениями [797, 861, 862].
Анализ литературных данных, характеризующих поведенческие реакции рыб в токсических растворах, позволяет предполагать, что их использование в качестве характерных симптомов отравления той или иной группой ядов пока затруднительно, поскольку эти симптомы могут быть различными у разных видов рыб при действии одного и того же вещества и сходными при отравлении их разными веществами. Достаточно сказать, что такие поведенческие реакции рыб на токсические воздействия, как двигательное возбуждение, потеря рефлекса равновесия, опрокидывание на бок, угнетение дыхания и т. д., могут иметь место при отравлении аммиаком, цианидами, сульфидами, этиловым спиртом, различными органическими ядами и даже некоторыми тяжелыми металлами. Это не значит, конечно, что поведение рыб не имеет диагностического значения.
Напротив, поведенческие реакции, их изменение являются наиболее ранними и чувствительными тестами отравления рыб сублетальными концентрациями различных токсикантов и потому могут с успехом использоваться для определения самого факта токсичности данной группы веществ. Другое дело, если авторы пытаются использовать поведенческие реакции рыб для дифференциальной диагностики отравления, т. е. для выявления вида вещества или группы веществ, вызвавших отравление данного вида, рыб в естественном водоеме или в экспериментальных условиях. В данном случае опыты с использованием одного вида рыб и ограниченного набора токсикантов могут ввести в заблуждение. Именно на основе таких опытов чаще всего судят о специфичности поведенческих симптомов отравления тем или иным токсикантом.
Показательны в этом отношении опыты [366], проведенные на культурном карпе, в которых регистрировались симптомы острого отравления четырьмя веществами: фенолом, полихлорпиненом, хлорофосом и одним из детергентов («Лотос-71»). На основе полученных данных автор приходит к выводу, что «при воздействии полихлорпинена в отличие от фенола у рыб во время бурной двигательной активности наблюдается переход в поверхностный слой и такой специфический симптом, как заглатывание воздуха. При отравлении хлорофосом характерны длительная стадия торможения, а также из-за сильного раскрытия хроматофор потемнение окраски. При действии детергентов отмечается обильное выделение слизи.
Еще более специфичны симптомы отравления у одного из представителей беспозвоночных — медицинской пиявки в растворах вышеупомянутых веществ [366, с. 268]. Возможно, применительно к карпу такая специфика поведенческих реакций имеет место, однако если рассмотреть эти материалы в свете литературных данных, характеризующих поведенческие реакции различных видов рыб, отравленных, скажем, тем же фенолом, то неизбежно приходишь к выводу об отсутствии такой специфики. Например, при отравлении угрей [469] и налимов [163] фенолом выявляется длительная стадия торможения, а у некоторых видов рыб из-за раскрытия хроматофор — потемнение окраски спины и головы [163]. Ясно, что эти же симптомы восприняты как специфические у карпов, отравленных хлорофосом. Далее, обильное выделение слизи характерно якобы при действии детергентов на карпа, но этот же признак указывается в качестве специфического показателя фенольного отравления рыб [594, 831, 432, 775].
Безусловно, поведенческие реакции могут использоваться в качестве первоначальных признаков нарушения нормальной жизнедеятельности отравленных рыб, но выявить специфические компоненты этих реакций, характерные для той или иной группы веществ, а тем более отдельного вещества, — задача чрезвычайно трудная. Эта трудность определяется зависимостью поведенческих реакций от вида рыб, химических свойств токсиканта и его концентрации, что значительно усложняет использование поведенческих реакций для дифференциальной диагностики отравления рыб.
Более реальную помощь ихтиотоксикологам может оказать, как нам представляется, патоморфологическое и гистопатологическое изучение картины отравления тем или иным токсикантом. Знание изменений в органах и тканях рыб может быть использовано для выявления путей проникновения токсикантов и их локализации в организме, понимания патогенеза и прогнозирования исхода токсического процесса, а также для дифференциальной диагностики токсикозов рыб, что имеет уже непосредственное практическое значение для выявления причин гибели рыб в естественных водоемах.
Первые опыты в этом направлении обнадеживали. По данным А. О. Таусон [338], различные группы веществ оказывают неодинаковое воздействие на покров тела подопытных рыб. Некоторые органические вещества (анилин, нафтеновые кислоты) вызывают свертывание слизи и ее отслаивание от тела в виде хлопьевидных образований, а сильные минеральные кислоты (серная и соляная) — также свертывание слизи, но с последующей фиксацией и помутнением покровов тела. Соли щелочноземельных металлов обладают стимулирующим действием на секрецию слизи. Патологоанатомическое изучение гольянов, отравленных нитритом натрия, выявило [641] увеличенное отложение продуктов распада гемоглобина в печени, селезенке и почках, а также анемию.
Специфические морфологические изменения обнаружены при отравлении карпа и радужной форели фтористым натрием [725] и большеротого черного окуня шестивалентным хромом [554]. М. М. Телитченко и Е. Н. Левитова [347] показали, что гистологический анализ гонад позволяет вскрыть ранние симптомы отравления ураном-289 и обнаружить даже залповые сбросы токсических веществ в водоем. Длительное воздействие азотнокислого уранила на уклеек (Alburnus alburnus-L.) уже в концентрациях 1, 5, 10 и 25 мг/л вызывало дегенерацию яичников или их перерождение в семенники. Сходные изменения, т. е. маскулинизация самок рыб, происходит также под влиянием таких факторов, как повышенная температура, свет, содержание в малых аквариумах в период интенсивного роста, и, следовательно, носят неспецифический характер.
Детальное гистопатологическое изучение острого отравления рыб водными экстрактами продуктов каменноугольной смолы, водорастворимыми компонентами сырой нефти, мазутом и фенолом позволило выявить [51] общие и специфические черты отравления рыб (сазан, золотая рыбка) этими веществами. Патоморфологиче-ская картина отравления рыб водорастворимыми компонентами сырой нефти, каменноугольной смолы и мазута отличается от картины их отравления фенолом (разный характер поражения тканей и органов и их локализация). Первая группа ядов вызывает воспалительные и некробиотические изменения в жаберном аппарате и нижних отделах пищеварительного тракта, что не отмечалось при действии фенола на рыб. Во всех исследованных органах наблюдалось расстройство кровообращения с гиперемией и выходом эритроцитов за пределы сосудистого русла.
Общими признаками отравления ядами обеих групп были дистрофические изменения в паренхиматозных органах (сердце, печень, почки) с отложением желтого гемоглобинного пигмента. Помимо этих признаков у рыб, отравленных фенолом, отмечают коагуляцию крови в полости сердца и жабрах [588], но аналогичные изменения отмечены и при других воздействиях на рыб, в частности при асфиксии, и, следовательно, они не специфичны для фенольной интоксикации.
Весьма противоречивы сведения относительно характера изменений жаберного аппарата при фенольном отравлении рыб. По мнению Ф. Е. Вишневецкого [51], жаберный аппарат золотых рыбок, отравленных фенолом, почти не поражается, что согласуется с более ранними данными И. Гавелки и М. Эффенбергера [588]. Последующее обстоятельное изучение этого вопроса, выполненное на разных видах рыб X. Рейхенбах-Клинке [755, 756] и Д. Валютой [831—833], опровергло эту точку зрения. У рыб, отравленных фенолом, жаберные лепестки вздуты, гиперемированы и происходит отторжение покровных эпителиальных клеток, что свидетельствует о существенных изменениях жаберного аппарата. Особо следует отметить, что сходные изменения в строении жаберного аппарата отмечены у золотых рыбок, отравленных солями тяжелых металлов, кислотами и щелочами [776, 652, 575], а также у сомов, отравленных гептахло: ром и никотином [644, 645].
В наших опытах [54, 55], поставленных на икре и эмбрионах леща и синца, установлено, что токсический эффект фенола проявляется резким нарушением нормального хода эмбриогенеза и различными видами уродств. Наиболее распространенными были изогнутость туловища, а также нарушения в строении сердечно-сосудистой системы (водянка).
Сходный характер патоморфологических и патолого-анатомических изменений наблюдается и при других чрезвычайных по силе воздействиях на эмбриональное развитие рыб, в частности при резких колебаниях температуры.
Иными словами, выявленные нами патоморфологиче-ские изменения, развивающиеся при фенольном отравлении рыб, носят неспецифический характер и стереотипно воспроизводятся при многих стрессорных воздействиях на эмбриональное развитие рыб. Результаты этих опытов, а также критический анализ имевшихся к тому времени литературных данных позволили нам сделать вывод [163] о необходимости дальнейшего патоморфоло-гического и гистопатологического изучения симптомов отравления рыб для составления окончательного мнения о возможности использования морфологических показателей для дифференциальной диагностики токсикозов рыб в естественных водоемах.
За минувшие 15 лет интерес к этим вопросам заметно возрос как у нас в стране, так и за рубежом [432, 644, 645, 775, 541, 70, 407, 785, 124, 453, 786, 219]. Обстоятельное изучение гистопатологических изменений у леща при остром отравлении фенолом привело А. Г. Андреса и Т. Н. Куражковскую [9] к выводу о том, что «в картине отравления доминируют токсико-дегеиеративиые изменения органов, стоящих на пути циркуляции яда (жабры, кровеносная система, сердце, кишечный тракт, печень, почки). В высшей степени характерны также дегенеративные изменения всей мышечной системы (поперечнополосатой, скелетной и сердечной, а также гладкой мускулатуры кишечника). В целом гистопатология острого фенольного отравления представляет весьма типичный симптомокомплекс, резко от.рчный от пато-морфологии леща при дефиците кислорода или интоксикации продуктами каменноугольной смолы сазана» (с. 85).
Этот вывод был подвергнут обоснованной критике [216, 222]. По мнению В. Е. Матей [216], описанная авторами гистопатология отравления фенолом лещей не может считаться ярко специфичной, поскольку аналогичные изменения в паренхиматозных органах, стоящих на пути циркуляции яда, имеют место и при действии на рыб других токсикантов: диметилсульфооксида [447], ацетонитрила и диметилформамида [21, 22], ДДТ [638].
В повторных опытах Ф. И. Межнина [222], выполненных на модели острого фенольного отравления гуппи, особое внимание было сосредоточено на характере и выраженности гистопатологических изменений поперечнополосатых мышц, которые, по мнению А. Г. Андреса и Т. Н. Куражковской, «в высшей степени характерны для фенольного отравления рыб. На основе выполненного исследования Ф. И. Межнин считает, что поперечнополосатые мышцы в меньшей степени подвергаются изменениям при действии фенола, чем другие органы и; ткани. Критически оценивая экспериментальные данные А. Г. Андреса. и Т. Н. Куражковской, Ф. И. Межник [222] пишет: «Судя по рисункам, которыми авторы иллюстрируют изменения поперечнополосатых мышц при отравлении фенолом, они скорее отражают артефакты, возникающие при гистологической обработке материала, чем патологические изменения мышц» (с. 51). К этому следует добавить, что и с физиологических позиций мнение А. Г. Андреса и Т. Н. Куражковской [9] о том, что «ярко представленные в клинической картине этой интоксикации рыб двигательные расстройства с терминальными параличами имеют не только нервный генез..., но и связаны с непосредственными повреждениями мышечного аппарата» (с. 84), вызывает серьезные возражения. Экспериментальные исследования, выполненные на спииальных карасях, а также на карасях, у которых полностью вылущен головной мозг, показали, что фенол не оказывает прямого раздражающего действия на туловищные мышцы рыб [158, 163] и, следовательно, двигательные расстройства с терминальными параличами имеют центральное, а не периферическое происхождение.
Детальный критический анализ работы А. Г. Андреса и Т. Н. Куражковской [9], утверждающей специфичность морфологических изменений у рыб, на основе опытов с одним токсикантом и на одном виде рыб в полной мере вскрывает несостоятельность такого методологического подхода и вновь указывает на необходимость привлечения литературных сведений при обсуждении собственных экспериментальных данных, иначе ошибочные выводы неизбежны. Этим строгим требованиям в полной мере отвечает исследование Ф. И. Межнина [223], направленное на выявление патогистологических изменений органов и тканей гуппи при остром отравлении фенолом, полихлорпиненом и повышенной солевой нагрузке.
Сопоставление результатов собственных исследований с литературными данными приводит автора к выводу, что морфологическая картина поврежденных органов оказывается примерно одинаковой у рыб, отравленных различными токсикантами, что, естественно, затрудняет использование морфологических симптомов для дифференциальной диагностики токсикозов.
Анализ конкретных экспериментальных данных, характеризующих степень поражения различных органов и тканей рыб отдельными токсикантами, показывает, что структура одних органов (жабры, почки, печень) претерпевает у отравленных рыб более существенные изменения, чем структура других (поперечнополосатые мышцы, сердце, мозг). Наиболее глубокие изменения отмечаются в органах, стоящих на пути циркуляции токсикантов, например в жабрах. Они первыми подвергаются действию токсикантов, нарушающему структуру этого жизненно важного органа дыхания и выделения. При этом в первую очередь слущивается эпителий жабр либо полностью (полихлорпинен), либо настолько, что у основания жаберных лепестков остаются лишь отдельные эпителиальные клетки (фенол, поваренная соль). Одновременно происходит переполнение кровеносной системы жабр).
Аналогичные изменения в жабрах рыб имеют место при дефиците кислорода [8], отравлении рыб фенолом [755, 831, 9, 222], додецилбензолсудьфонатом-ДБС; гептахлором [645], 2-метил-5-этилпиридином-МЭП и 2-метил-5-винилпиридином-МВП [383], а также сульфатом меди [432], мазутом, нефтью и каменноугольной смолой [51]. Следовательно, при отравлении рыб токсикантами различной природы, а также при повышенной солевой нагрузке и дефиците кислорода характер поражения жабр одинаков, различна лишь степень регистрируемых структурных изменений. На основе этих данных Ф. И. Межнин [223] приходит к вполне обоснованному выводу о том, что «о специфичности поражения жабр при токсикозах рыб говорить не приходится, и картина жабр не может являться дифференциальным признаком (при указанных токсикантах) в диагностике токсикозов рыб» (с. 60).
Сказанное в полной мере относится и к морфологическим изменениям сердца у отравленных рыб. В ответ на воздействие различных токсикантов, таких, как фосфор [203], МЭП и МВП [383], фенол [9, 222], полихлорпинен [223], а также при повышенных солевых нагрузках [223] и дефиците кислорода [8] происходит гипертрофия желудочка и инфильтрация лимфоцитами. Следовательно, характер поражения сердца при различных токсикозах также не может быть надежным основанием для дифференциальной диагностики отравления рыбы тем или иным токсикантом.
Сходные патологические изменения отмечены и в почках рыб, отравленных различными токсикантами. Большинство исследователей описывают расстройство кровообращения, различную степень деструкции почечных клубочков и канальцев, активацию эритропоэза. Последняя реакция носит защитный, а точнее, компенсаторный характер. Дело в том, что при многих токсикозах рыбы испытывают недостаток кислорода. Для обеспечения организма кислородом в этих стрессорных ситуациях на первом этапе происходят выброс эритроцитов из «депо» в кровяное русло и переполнение кровеносной системы внутренних органов, а также головного и спинного мозга [755; 831, 9, 383, 223]. Значительные морфологические изменения, развивающиеся в мозге рыб под действием различных токсикантов, — гиперемия мозга, сжатие тканевых элементов, наличие больших областей вакуолизации [638, 22, 216] — также имеют неспецифический характер. Вслед за первой фазой - компенсаторной реакции в ответ на возникающий в организме отравленных рыб дефицит кислорода — выход запасов крови в кровеносное русло, развивается вторая фаза — усиление эритропоэза в почках.
Еще один важнейший внутренний орган — печень — в ответ на токсическое воздействие различной природы реагирует также однотипно. Здесь развивается жировая дистрофия, происходит переполнение сосудов кровью [223], сопровождаемые снижением включения гликогена в печеночных клетках [382, 383] и очаговой инфильтрацией лимфоидными элементами [203].
Обобщая заслуживающие внимания экспериментальные данные, можно считать доказанным, что патоморфологические изменения в различных органах рыб, вызванные разными по своей природе токсикантами, весьма сходны и потому «патогистологическая картина при отравлениях рыб вряд ли может иметь решающее значение в дифференциальной диагностике токсикозов рыб» [223, с. 64]. Большие надежды, возлагавшиеся на патоморфологические исследования применительно к диагностике отравлений рыб [202], не оправдались. Однако это не значит, что продолжать подобного рода исследования нецелесообразно. Многие теоретические и практические вопросы ихтиотоксикологии могут быть решены только при комплексном изучении реакций рыб на различные группы токсикантов с использованием биохимических, физиологических и гистологических методов исследований. Особое значение патоморфологические и гистопатологические исследования приобретают при разработке проблемы аккумуляции различных токсикантов и в первую очередь стойких пестицидов в органах и тканях рыб, оценке степени вредности различных уровней накопления вредных веществ при их длительном нахождении в организме, выявлении органа-мишени для различных групп токсикантов, понимании течения и сущности патологических процессов. Объективная регистрация картины поражений важнейших органов при действии различных групп токсикантов у разных видов рыб может оказать существенную помощь в раскрытии механизмов развития острого н хронического отравления.








