 2020-05-12
2020-05-12 278
278
|
|

Высокая скорость обратимости токсического процесса у рыб отмечается, как мы уже упоминали выше, и другими авторами [628, 596]. Обращает внимание однотипная динамика обратимости фенольной интоксикации при всех изученных нами концентрациях. Восстановление различных рефлексов идет в обратном порядке сравнительно с прямым развитием фенольной интоксикации. Функции, нарушенные в первую очередь, восстанавливаются последними.
Караси, находящиеся в боковом положении, сопровождающемся конвульсивными судорогами (третья фаза), постепенно приобретают способность сперва импульсивного, а затем и более плавного перемещения в боковом положении (вторая фаза), и, наконец, рефлекс равновесия полностью восстанавливается. Таким образом, процесс восстановления функций также проходит последовательно три фазы, описанные нами ранее.
Сопоставляя степень обратимости различных фаз фенольной интоксикации, необходимо отметить, что первая фаза — фаза резкой двигательной возбудимости — во всех случаях обратима. Вторая фаза фенольной интоксикации, объединяющая такие реакции рыб, как потеря рефлекса равновесия, опрокидывание на бок и импульсивное перемещение в боковом положении, также высокообратима даже в растворах фенола концентрацией 200 и 400 мг/л. Что касается третьей фазы фенольной интоксикации, характеризующейся потерей двигательной активности и конвульсивными судорогами в боковом положении, то ее обратимость значительно ниже. Отмечаются случаи полной обратимости интоксикации (в низких концентрациях фенола), частичной ее обратимости без восстановления рефлекса равновесия и полной необратимости интоксикации с последующей гибелью испытуемых рыб, хотя и в более отдаленные сроки.
Кривая, характеризующая степень обратимости интоксикации у рыб, вызванной различными концентрациями фенола, показана на рис. 3. Характеризуя особенности обратимости интоксикации рыб в растворах с различней концентрацией фенола, можно отметить, что после нахождения рыб на протяжении 15,5 ч в токсическом растворе (50*мг/л) имела место 100%-ная обратимость интоксикации. Рыбы были перемещены в чистую воду во второй фазе развития фенольной интоксикации. При 5,5-часовой экспозиции испытуемых рыб в растворе концентрацией 100 мг/л большинство рыб было перенесено в чистую воду в третьей фазе развития интоксикации. Как видно из рис. 3, 70 % рыб выжили и 30 % погибли. При этом рефлекс равновесия быстро восстанавливался у 4 из 7 рыб, а 1 из 3 других рыб находилась в боковом положении на протяжении всего срока наблюдений. (10 сут).
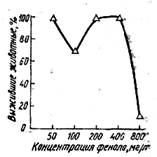
Рис. 3. Степень обратимости фенольной интоксикации карасей
Степень и динамика обратимости интоксикации карасей в растворах фенола концентрацией 200 и 400 мг/л имели примерно сходный характер. Среднее время экспозиции рыб в растворах было также приблизительно одинаковым (8,3 и 6,4 ч соответственно). Рыбы при перемещении из растворов в чистую воду находились частично во второй и частично в третьей фазе развития фенольной интоксикации. В растворе фенола концентрацией 200 мг/л восстановление рефлекса равновесия отмечено у всех рыб и в сравнительно короткий срок (24 ч). В растворе фенола концентрацией 400 мг/л полная обратимость у 9 рыб была достигнута за 24-26 ч, а 1 рыба находилась в боковом положении до конца наблюдений. Концентрация фенола 800 мг/л в течение 2 л вызывала необратимую интоксикацию в 90% случаев, и только одна рыба из 10 оставалась живой, у нее полностью восстанавливался рефлекс равновесия после перенесения в чистую воду. В этом растворе фенола время гибели 50% испытуемых рыб мало отличается или почти не отличается от средней продолжительности выживания всей выборочной совокупности.
Полученные экспериментальные данные по динамике обратимости фенольной интоксикации позволяют сделать вывод о высокой скорости и степени обратимости фенольного отравления рыб. При этом трудно отказаться от мнения, что время нахождения испытуемых рыб в токсическом растворе играет более существенную роль, чем концентрация яда. Это подтверждается примерно одинаковой обратимостью интоксикации в диапазоне концентраций фенола от 50 до 400 мг/л.
Последующими нашими опытами, проведенными с фенолом и тремя изомерами крезола на плотве, густере и язе, в общих чертах были подтверждены результаты опытов на карасях. Вместе с тем степень обратимости интоксикации зависит как от видовых особенностей испытуемых рыб, так и от химических свойств исследуемого вещества. Так, например, оказалось, что фенольная интоксикация плотвы более легко обратима, чем крезольная, а интоксикация у густеры менее обратима, чем у плотвы.
Большой практический интерес представляет вопрос об обратимости интоксикаций рыб, вызванных действием инсектицидов. Согласно опытам X. Манна [689] плотва, отравленная фост-фитоном или тиоданом (30 мин при концентрации 0,01 мг/л), приходила в нормальное состояние после перемещения в чистую воду, однако действие более высоких концентрация яда было необратимым. Д. Людеман и X. Нейман [678] провели сравнительное изучение токсичности 16 хлор- и фосфорорганических инсектицидов на карпах, щуках и форели и установили, что для хлорорганических соединений характерна необратимость действия, тогда как при отравлении фосфорорганическими инсектицидами рыбы, перемещенные в чистую воду, вскоре приходили в нормальное состояние.
Согласно представленным данным токсическое действие многих изученных веществ обратимо в случае прекращения контакта рыб с ядом. Степень обратимости токсического процесса в значительной мере определяется видовыми особенностями подопытных рыб и химическими свойствами испытуемого вещества. Однако большинство авторов не указывают длительность наблюдения за рыбами после их перемещения в чистую воду, а это может иметь решающее значение. Скорость восстановления рефлекса равновесия, нормализации дыхания и восстановление у рыб способности к плаванию (эти признаки чаще всего используются в качестве показателей обратимости токсического процесса) сами по себе не могут характеризовать нормализацию физиологического состояния и лежащих в его основе биохимических процессов у рыб, перенесших отравление. Для решения этого вопроса необходимо использование более тонких биохимических и физиологических показателей, отражающих состояние различных функциональных систем организма [163]. Последующие экспериментальные исследования полностью подтвердили высказанное предположение.
Серия опытов по обратимости токсического процесса у гуппи, отравленных фенолом, выполнена В. Е. Матей [216]. Показателем восстановления нормального состояния рыб, перенесших отравление, служили уровень условнорефлекторной деятельности и структурные изменения в гипоталамо-гипофизарной нейросекреторной системе. Предварительными опытами установлено, что наиболее значительные изменения высшей нервной деятельности у подопытных гуппи происходили под влиянием двух концентраций фенола: 6,25 и 12,5 мг/л. Условные реакции у подопытных рыб были либо резко угнетены (первая концентрация), либо их вообще не удавалось выработать (вторая концентрация). После перемещения подопытных гуппи из раствора фенола концентрацией 6,25 мг/л в чистую воду в первые 15 сут не отмечались какие-либо существенные изменения в их высшей нервной деятельности: латентные периоды реакций оставались высокими, процент проявления положительных условных рефлексов был низким, дифференцировочные условные рефлексы угнетены. Только на 38-е сутки пребывания рыб в чистой воде средние показатели условнорефлекторной деятельности у этих рыб становились выше, чем у рыб, находившихся в токсическом растворе, но были значительно ниже, чем у рыб, не подвергавшихся отравлению. В наименьшей степени восстанавливались дифференцировочные рефлексы, т. е. наиболее сложные формы условнорефлекторной деятельности. В еще более поздние сроки происходило частичное восстановление условнорефлекторной деятельности у рыб, перенесших более тяжелую форму фенольного отравления (концентрация 12,5 мг/л). У них только к исходу 2-го месяца нахождения в чистой воде (53-и сутки) появлялась принципиальная возможность выработки условных рефлексов.
Трудность полного восстановления функционального состояния отравленных рыб, долгое время находившихся в растворах фенола, подтвердилась и на примере гипота-ламо-гипофизарной нейросекреторной системы (ГТНС). Длительное воздействие (до 1 мес) высоких концентраций фенола (16,7 и 25 мг/л) вызывает ряд морфофункциональных изменений этой системы у подопытных гуппи: снижение, а затем и полное опустошение нейросекреторной системы от нейросекреторного вещества, сильное обводнение тканей мозга и гипофиза. Только после месячного нахождения отравленных рыб в чистой воде в преоптическом ядре гуппи повышалось содержание нейросекреторного вещества, патоморфологические изменения тканей мозга и гипофиза ослабевали, снижалась их гиперемия. Уровень функциональной активности у нейрогипофиза был выше, чем у нейросекреторных центров гипоталамуса. Степень восстановления патологических изменений в ГГНС находилась в обратной зависимости от концентрации токсического вещества. На основе полученных данных В. Е. Матей приходит к вполне обоснованному выводу о чрезвычайно сильном токсическом действии фенола на рыб с длительным периодом последействия.
Длительное последействие характерно не только для фенола, но и для других ядов органического ряда, в частности для пестицидов различного химического состава. Об этом свидетельствуют результаты обстоятельного исследования, выполненного в лаборатории М. М. Телитченко Мохамедом Али [3] на модели пестицидного отравления карпа.
Автор использовал в своих опытах 7 пестицидов, три из которых принадлежат к группе фосфорорганических (фосфамид, цидиал, дисистон), два — к группе хлорорганических (линдан и эндрин) и два — к группе карбаматных пестицидов (севин и темик). Показателем токсичности исследованных пестицидов служило угнетение активности ацетилхолинэстеразы мозга отравленных рыб, а показателем обратимости пестицидного отравления — возвращение уровня активности ацетилхолинэстеразы мозга к норме.
После 6-часового контакта с пестицидами разных концентраций рыб помещали в чистую воду. В течение первых суток наблюдения активность ацетилхолинэстеразы мозга продолжала снижаться, как и у рыб, находившихся в токсическом растворе. Восстановление активности фермента обычно начиналось через 2-6 сут после перевода рыб в чистую воду. Скорость и степень этого восстановления зависели от свойств пестицида и его концентрации, причем отмечена парадоксальная зависимость: у рыб, подвергавшихся действию более высоких концентраций пестицидов, восстановление начиналось раньше (через 1 сут), чем у рыб, испытавших действие низких концентраций этих же пестицидов. Например, в опытах с дисистоном концентрацией 5 мг/л восстановление активности фермента у отравленных рыб начиналось через 1 сут после перевода их в чистую воду, а у рыб, испытавших действие пестицида концентрацией 0,01 и 0,1 мг/л, — только через 6 сут.
При отравлении рыб малыми концентрациями пестицидов восстановление активности ацетилхолинэстеразы оказалось относительно меньшим, и к концу трехнедельных наблюдений последействие испытанных концентраций пестицидов было четко заметно. Степень обратимости действия разных пестицидов также неодинакова: через 3 нед. после погружения рыб в чистую воду активность фермента мозга восстанавливалась на 90%. У карпов, отравленных темиком, на 86% — отравленных дисистоном, до 75% — севином и лишь до 66% у рыб, отравленных фосфамидом.
Эти материалы хорошо согласуются с более ранними наблюдениями С. Вейса [842], изучавшего степень обратимости действия фосфорорганических пестицидов и установившего, что она зависит как от свойств пестицида, так и от вида рыб.
Так, например, 12-24-часовое выдерживание ушастого окуня (Lepomis macrochirus) и черноголовой пимефалес (Pimephales promeles) в растворе карбофоса концентрацией 0,1 мг/л приводило к снижению активности фермента на 82% у первого вида и только на 20% у второго. Восстановление активности ацетилхолинэстеразы у черноголовой пимефалес произошло через 8 сут, а у ушастого окуня — только через 30 сут. Пониженная устойчивость к интоксикации у ушастого окуня в сравнении с устойчивостью черноголовой пнмефалес, а также нотомигонуса (Notemigonus crysoleucas) и серебряного карася (Carassius auratus) отмечена автором в опытах с другими пестицидами (паратион, дельнав, ДДВФ). Видовые различия устойчивости к интоксикации трех видов лососевых рыб обнаружены в опытах с одним из широко применяемых фосфор-органических пестицидов — карбофосом [748]. 7-10-суточное выдерживание рыб в растворе карбофоса концентрацией 0,4-3,0 мг/л вызвало снижение активности ацетилхолинэстеразы мозга на 25% У ручьевой форели (Salvelinus foutinalis), на 45% У радужной форели (Salmo gairduerii) и на 75 % у кижуча (Oncorhynchus kisutsch). Полное восстановление активности фермента происходило через 25-42 сут.
Сходные данные получены Н. Ф. Косиновой [135], изучавшей обратимость токсического действия метафаса на карпе с использованием в качестве показателя восстановления физиологического состояния отравленных рыб активности ацетилхолинэстеразы в их органах и тканях, а также количества свободных SH-rpynn и активности пероксидазы.
Так, у рыб, переведенных из раствора метафоса (0,01 мг/л) в чистую воду, активность ацетилхолинэстеразы печени и мышц продолжает снижаться и на 5-е сутки составляет 65% контроля. Однако в дальнейшем отмечается постепенное повышение активности фермента до уровня контрольной группы (через 20—25 сут). В мозге активность ацетилхолинэстеразы нормализуется через 30 сут после перевода отравленных рыб в чистую воду, а в жабрах активность фермента хотя и увеличивается в сравнении с его активностью у отравленных рыб, но остается ниже активности фермента контрольных рыб в течение 25 сут пребывания в чистой воде.
Нормализация активности другого фермента — пероксидазы — в мышцах, жабрах и мозге рыб, а также количества тиоловых групп отмечается через 20- 25 сут. Полное восстановление исследованных биохимических показателей наступает через 30-45 сут после снятия токсического эффекта метафоса и совместного действия метафоса со стронцием-90.
Длительный период восстановления нормального физиологического состояния рыб, перенесших отравление, автор связывает с нахождением в их организме остаточных количеств инкорпорированных полюталтов и постепенной заменой у рыб, перенесших отравление метафосом, фосфорилированной холинэстеразы, гидролизующейся значительно медленнее ацетилированной, за счет усиленного синтеза нормального фермента в печени.
Несмотря на то, что в литературе отсутствуют сведения относительно скорости обновления белков печени у рыб, можно согласиться с предположением Н. Ф. Косиновой [135] о наличии причинно-следственной связи между усиленным синтезом в печени нормальной, неингибированной холинэстеразы и временем восстановления жизнедеятельности рыб, перенесших отравление, если допустить, что эта скорость обновления белков печени незначительно отличается от той, что имеет место у теплокровных животных, у которых этот процесс протекает с периодичностью в 14-20 сут [303]. Но, это лишь один из механизмов обратимости отравления рыб пестицидами, другим механизмом является усиленное выведение токсиканта из организма, которое по времени совпадает с частичным или полным восстановлением нормальной жизнедеятельности отравленных рыб.
Так, через 26 сут после перевода вьюнов из раствора ДДТ концентрацией 0,1 мг/л (3-суточная экспозиция) в чистую воду содержание п, n' -ДДД в организме рыб снизилось с 2,5-3,0 до 0,7-1,2 мкг на 1 г живой массы. Сходная по интенсивности, но более замедленная элиминация токсиканта отмечена и у карпа [267]. За 39 сут нахождения рыб в чистой воде содержание о, n' -ДДД в мышцах снизилось в 2-3 раза (с 6-8 до 3 и с 4,0 до 1,0-1,5 мкг на 1 г живой массы).
Обобщая добытые к настоящему времени экспериментальные данные, мы вновь, но уже на примере биохимических показателей, убеждаемся в длительном последействии токсического влияния различных групп веществ, поступающих в рыбохозяйственные водоемы. Это подтверждает высказанное ранее мнение [163] о необходимости более длительных наблюдений за рыбами, подвергнутыми действию токсикантов. Согласно нашим данным [189], полученным на сеголетках карася, из 16 выживших рыб в растворе фенола концентрацией 200 мг/л 9 погибли от сапролегнии в течение 4 последующих суток (или позже). В опытах на плотве и густере, отравленных фенолом или крезолом, мы имели возможность отметить при длительном наблюдении (до 28 сут) гибель части выживших после отравления рыб от сапролегнии. Следовательно, перенесенное рыбами отравление не проходит бесследно и может снижать их устойчивость к другим патогенным воздействиям, в частности грибковым или инфекционным заболеваниям.
Это дает возможность с новых позиций рассмотреть чрезвычайно важный в практическом отношении вопрос. Нередко приходится производить экспертизы массовой гибели рыб в том или ином водоеме, причины которой недостаточно ясны. По-видимому, вспышки заболеваний рыб имеют в своей основе хроническое отравление их малыми концентрациями ядов промышленных сточных вод или пестицидов, проявляющееся при ухудшении кислородного режима, изменении активной реакции среды, перепаде температур и других неблагоприятных условиях. Рыбы, ослабленные хроническим отравлением (различными токсикантами, легче подвергаются инфекционным и паразитарным заболеваниям вследствие снижения общей сопротивляемости организма [664, 189].
За последние 15 лет появились новые факты, раскрывающие различные стороны проблемы обратимости интоксикации рыб и ее значение для теории и практики ихтиотоксикологии. Действие многих токсических ве-Тцеств на рыб и, в частности ядов органического ряда, обратимо. Особое внимание исследователей привлекают различные группы пестицидов. В опытах с монуроном [398], форфамидом и метилнитрофосом [230, 291], малатионом [748], ДДТ [844, 267] отмечена способность рыб восстанавливать жизнедеятельность после снятия токсического действия и перевода их в чистую воду.
Вместе с тем накапливаются материалы, свидетельствующие о том, что отравление некоторыми веществами (помимо уже отмеченных — концентрированные растворы большинства минеральных кислот, соли тяжелых металлов, некоторые яды органического ряда) труднообратимо или вообще необратимо [53, 32, 128, 23].
Например, двухнедельное нахождение бычков и гольянов в сточной воде БЦЗ 10-, 20- и 30-кратного разведения приводит к необратимым изменениям гематологических показателей [53]. Длительное (30-суточное) пребывание рыб в отравленном монуроксом водоеме вызывает необратимые патологические изменения форменных элементов крови [128]. За 45 сут нахождения в чистой воде погибло около 70% отравленных рыб. Высокий процент гибели отравленных ацетонитрилом лещей отмечен Ю. А. Белоусовым [23] при длительном (60-90 сут) контакте рыб с ядом. При 60-суточной экспозиции лещей в растворе ацетонитрила концентрацией 10 мг/л обратимость интоксикации после 10-суточного нахождения в чистой воде составила лишь 60%, а при 90-суточной экспозиции все рыбы, перенесенные в чистую воду, погибли на 2-е сутки. Сокращение времени экспозиции лещей в токсическом растворе до 30 и 15 сут значительно увеличивало обратимость интоксикации.
Результаты этих опытов в совокупности с полученными нами ранее данными [189, 163] показывают, что степень обратимости той или иной интоксикации зависит от концентрации токсиканта и времени нахождения рыб в токсическом растворе. Необходимы дальнейшие целенаправленные поиски отдаленных последствий перенесенного рыбами отравления (острого или подострого) и их влияния на устойчивость рыб к стрессорным воздействиям различной природы. Для более полного ответа на вопрос о степени обратимости отравления рыб тем или иным токсикантом следует увеличить время наблюдения за рыбами, перенесшими отравление, от 2 до 4 иед, а в конце опытов весьма желательно оценить полноту восстановления нормального состояния методом функциональных нагрузок [163, 174].








