 2020-05-25
2020-05-25 157
157Эффективность развития фолликулов в яичниках зависит от регулирующего влияния на них гонадотропинов аденогипофиза. Рецепторы, на мембране клеток внутренней оболочки — к лютропину (РЛт) и на мембране гранулезных клеток — к фоллитропину (РФт), появляются в преантральной и ранней антральной стадиях развития фолликула. Связывание гормонов с рецепторами к лютропину и фоллитропину стимулирует в клетках синтез стероидных гормонов. Лютропин в текальных клетках стимулирует превращение ацетата и холестерина в андрогены (70 % от циркулирующих в плазме крови). Под влиянием лютропина текальные клетки синтезируют небольшое количество эстрогенов. Эстрогены и андрогены в небольшом количестве диффундируют в гранулезные клетки и под влиянием фоллитропина эти гормоны ароматизируются в эстрогены. Основными эстрогенами являются эстрадиол-170 и эстрон.
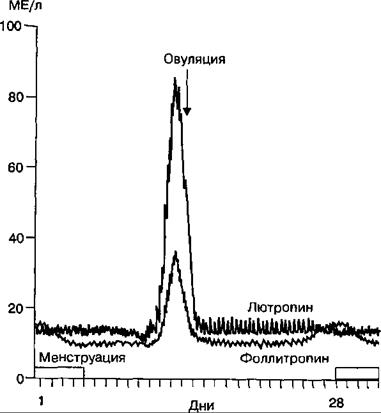
Рис. 16.10. Динамика содержания лютропина и фоллитропина в плазме крови в течение менструального цикла.
Успешное развитие овуляции во время короткой овуляторной фазы зависит от резкого и быстро проходящего пика секреции лютропина и фоллитропина. Пик секреции гонадотропинов нарастает в течение примерно 15 ч, достигает относительно постоянного уровня (следующие 15 ч), а затем снижается до исходного уровня в течение последующих 20 ч.
ной фазы один из фолликулов, на мембране которого выше, чем у других фолликулов, плотность рецепторов к фоллитропину, достигает размеров в среднем 11 мм и становится доминантным (вторичный фолликул). Это обусловлено тем, что в нем синтезируется больше, чем в других фолликулах, эстрадиола-17р. При высокой плотности рецепторов к фоллитропину на мембране доминантного фолликула, в нем сохраняется способность синтезировать эстрадиол-17р во время циклического снижения секреции фоллитропина в аденогипофизе женщины. В этих условиях другие фолликулы, имеющие низкую плотность рецепторов к фоллитропину на своей мембране, синтезируют незначительное количество эстрадиола-17р и подвергаются атрезии. Функция фоллитропина в развитии доминантного фолликула заключается в следующем. Этот гормон связывается с рецепторами мембраны гранулезных клеток и стимулирует в них синтез ароматазы, превращающей тестостерон в эстрадиол^ 17р. Тестостерон синтезируется во внутренних клетках наружной оболочки фолликула (theca) и диффундирует в гранулезные клетки, где гормон превращается в эстрогены (рис. 16.9). Количество андрогенов фолликулярного происхождения у женщины составляет примерно 70 % от их общей концентрации в плазме крови. Гранулезные клетки содержат эстрогеновые рецепторы, с которыми связывается образующийся в них эстрадиол и стимулирует пролиферацию этих клеток, увеличивая размер фолликула. Одновременно эстрадиол-17(3 через р-тип эстрогенных рецепторов активирует в гранулезных клетках абразова- ние новых рецепторов к фоллитропину. Поэтому чем больше образуется гранулезных клеток в фолликуле, тем больше андрогенов ароматизируется в эстрогены, которые стимулируют продукцию гранулезными клетками еще большего количества эстрадиола-17р (механизм положительной обратной связи). Нарастающий синтез эстрогенов в гранулезных клетках приводит к повышению концентрации женских половых гормонов в плазме крови. Эстрогены вместе с фоллитропицом стимулируют образование на мембране гранулезных клеток рецепторов к лютропину, который является основным регулятором следующей стадии развития антрального фолликула (третичный фолликул). Лютропин связывается с рецепторами на мембране гранулезных клеток и клеток внутреннего слоя наружной мембраны (theca) и стимулирует накопление в них липидов, желтого пигмента, а также пре- овуляторное увеличение образования прогестерона, который инициирует овуляцию. В конце фолликулярной фазы под влиянием нарастающей секреции эстрогенов в фолликуле и увеличения в плазме крови концентрации яичникового цитокина ингибина, тормозится секреция фоллитропина в аденогипофизе по механизму отрицательной обратной связи. При этом высокий уровень эстрогенов в плазме крови вызывает торможение секреции гонадолиберина в гипоталамусе и фоллитропина в гипофизе. Напротив, эти же гормоны (эстрогены и ингибин) по механизму положительной обратной связи стимулируют резкое повышение концентрации лютропина в плазме крови за 24—36 ч до начала овуляции (рис. 16.10). Нормальная овуляция во время короткой овуляторной фазы зависит от выброса, в виде быстро проходящего пика концентрации в плазме крови гонадотропинов, прежде всего лютропина.
16.3.1.2.Овуляторная фаза
Овуляция происходит примерно через 15 ч после достижения концентрацией лютропина в плазме крови максимальных значений (пик концентрации). Лютропин связывается с рецепторами мембраны фолликулярных клеток и активирует завершение первого мейотического деления первичного ооцита (см. рис. 16.1). В результате образуется вторичный ооцит, имеющий гаплоидное число хромосом, которые во время оплодотворения яйцеклетки сливаются с гаплоидным числом хромосом сперматозоида.
Кроме того, лютропин, связываясь с рецепторами системы G-белков фолликулярных клеток, увеличивает в них содержание вторичного посредника цАМФ. Под влиянием цАМФ в фолликулярных клетках непосредственно перед овуляцией прекращается фолликулогенез, связанный с образованием в гранулезных клетках стероидогенных энзимов, рецепторов фоллитропина, p-эстрогеновых рецепторов и циклина D2, являющегося регулятором митоза в фолликулярных клетках.
Под влиянием высокой концентрации лютропина в плазме крови возрастает кровоток в капиллярах ткани фолликула за счет роста продукции NO-синтазы и образования вазодилататора оксида азота в гладких мышцах сосудов, питающих фолликул. Это увеличивает размер фолликула, стимулируя накопление фолликулярной жидкости, которая своим гидростатическим давлением истончает слой гранулезных клеток и клеток наружной оболочки (theca) настолько, что лишь эпителиальный слой, покрывающий поверхность яичника, отделяет стенку фолликула от перитонеального пространства. В предовуляторную фазу лютропин стимулирует синтез рецепторов прогестерона, пептида, активирующего синтез аденилатциклазы в гипофизе, регуляторного фактора роста, которые вызывают овуляцию.
Преовуляторное увеличение концентрации прогестерона в плазме крови женщины активирует синтез в фолликуле протеолитических энзимов и простагландинов (Е и F). Под действием простагландинов, а также тромбоксанов и лейкотриенов происходит истончение, растяжение и разрушение стенки фолликула. В результате зрелая яйцеклетка выходит через стенку лопнувшего фолликула в жидкость перитонеального простраства. Двигательная активность цилий ампулярного расширения фаллопиевой трубы направляет движение ооцита в просвет фаллопиевой трубы.
16.3.1.3.Лютеальная фаза
После овуляции на месте лопнувшего фолликула образуется желтое тело, функцией которого является секреция эстрадиола-17(3, эстрона и прогестерона, регулирующих подготовку эндометрия матки к имплантации яйцеклетки. Если не происходит оплодотворения яйцеклетки, то желтое тело функционирует в лютеальную фазу овариального цикла (меструальное желтое тело). При оплодотворении яйцеклетки желтое тело функционирует первую половину беременности (желтое тело беременности), а затем его функция прекращается. Преовуляторное резкое повышение концентрации лютропина в плазме крови женщины приводит к лютеинизации гранулезных клеток и клеток theca и изменяет в них стероидогенез таким образом, что после овуляции в плазме крови из женских половых гормонов доминирует концентрация прогестерона. Связывание лютропина с рецепторами системы G-белков на мембране лютеальных клеток увеличивает содержание в них вторичных посредников, которые стимулируют транспорт холестерина из плазмы крови в клетки желтого тела, где он используется для синтеза прогестерона. Концентрация этого гормона в плазме крови женщины в лютеальную фазу зависит от массы стероидогенной ткани в желтом теле, интенсивности кровотока и способности стероидогенной ткани синтезировать прогестерон под влиянием лютропина. Стероидогенез в клетках желтого тела постепенно увеличивается и достигает пика примерно на 8—9-й день после овуляции. В этот период концентрация прогестерона в плазме крови женщины в 5 раз выше, чем в другие фазы овариального цикла. Повышение в плазме крови эстрогенов и прогестерона в лютеальную фазу по механизму отрицательной обратной связи тормозит секреции гонадотропинов в аденогипофизе, увеличивая с 1 до 2—3 ч интервал времени между периодами импульсного выброса этих гормонов кровь. В результате не начинается развитие очередного фолликула в яичниках. Секреция прогестерона в лютеальную фазу контролирует подготовку (секреторная фаза маточного цикла) эндометрия матки к имплантации яйцеклетки. В эту фазу происходит функциональное созревание клеток эндометрия, который полностью подготавливается к имплантации яйцеклетки. Если яйцеклетка не оплодотворяется, то желтое тело сохраняется около 14 дней и затем подвергается лютеолизу.
16.3.1.4.Лютеолиз желтого тела
Лизис, или структурное разрушение, желтого тела называется лютеолизом. В процессе лютеолиза утрачивается способность клеток желтого тела синтезировать и секретировать прогестерон. Физиологическое значение прекращения эндокринной функции желтого тела заключается в завершении овариального цикл, а также менструального цикла, поскольку не произошло оплодотворения яйцеклетки.
Лютеолиз регулируется простагландином ПГР2а, который образуется в клетках основной оболочки (theca) желтого тела. Простагландин ПГР2а связывается со специфическими метаботропными рецепторами, расположенными на мембране гранулезных стероидогенных клеток желтого тела. Взаимодействие простагландина ПГГ^ с рецептором мембраны стероидогенных клеток через систему G-белков и вторичные посредники инозитол- 3-фосфат и диацилглицерол активирует фосфолипазу С, которая вызывает в позднюю фазу развития желтого тела гидролиз мембранных фосфолипидов с высвобождением арахидоновой кислоты. Арахидоновая кислота, в свою очередь, является источником для синтеза простагландина ПГР2сх, что приводит к дополнительному росту концентрации простагландина и активации лютеолиза. Последний усиливается под влиянием эндотелина-1, секретируемого эндотелиальными клетками кровеносных сосудов желтого тела. Эндотелии-1 вызывает спазм сосудов желтого тела, прекращение в лютеальной ткани кровотока и, как следствие, разрушение стероидогенных клеток из-за недостатка в них кислорода и питательных веществ.
Удаление разрушенных клеток желтого тела осуществляется клетками крови (лейкоцитами, эозинофилами и макрофагами). Т-лимфоциты в желтом теле секретируют у-интерферон, с помощью которого иммунокомпетентные клетки распознают поврежденные клетки желтого тела и удаляют их из ткани яичника. Макрофаги фагоцитируют дегенеративные лютеальные клетки, тормозят с помощью цитокинов секрецию прогестерона в клетках желтого тела и активируют секрецию основного активатора лютеолиза простагландина ПГР2а. Макрофаги продуцируют a-фактор некроза опухоли, который тормозит секрецию прогестерона клетками желтого тела. Продукцию прогестерона ПГР2а активирует интерлейкин-I, который высвобождается макрофагами, фибробластами и эндотелиальными клетками капилляров желтого тела, что также усиливает лютеолиз. При участии иммунных клеток крови и цитокинов во время лютеолиза регулируется синтез простагландина ПГР2а в клетках желтого тела, в них тормозится стероидогенез и активируется фагоцитоз клеток в период его структурной инволюции.
16.3.2. Менструальный цикл (маточный цикл)
Подготовка женского организма к гестации характеризуется циклическими изменениями эндометрия матки, которые состоят из трех последовательных фаз: менструальной, пролиферативной и секреторной — и называются маточным, или менструальным, циклом.
16.3.2.1. Менструальная фаза
Менструальная фаза при длительности маточного цикла 28 дней продолжается в среднем 5 дней. Эта фаза представляет собой кровотечение из полости матки, которое возникает в конце овариального цикла, если не происходит фертилизации и имплантации яйцеклетки. Менструация — это процесс отторжения слоя эндометрия. Пролиферативная и секреторная фазы менструального цикла включают процессы восстановления эндометрия для возможной имплантации яйцеклетки в течение следующего овариального цикла.
16.3.2.2. Пролиферативная фаза
Пролиферативная фаза варьирует по продолжительности от 7 до 11 дней. Эта фаза совпадает с фолликулярной и овуляторной фазами яичникового цикла, в течение которого возрастает уровень эстрогенов, в основном эстрадиола- 170, в плазме крови. Основная функция эстрогенов в пролиферативную фазу менструального цикла — стимуляция клеточной пролиферации тканей органов репродуктивной системы с восстановлением функционального слоя эндометрия и развитием эпителиальной выстилки слизистой оболочки матки. В эту фазу под влиянием эстрогенов происходит утолщение эндометрия матки, увеличение в размере его желез, секретирующих слизь, растет длина спиральных артерий. Эстрогены вызывают пролиферацию эпителия влагалища, усиливают секрецию слизи в шейке матки. Секреция становится обильной, в ее составе увеличивается количество воды, что облегчает движение в ней сперматозоидов. Стимуляция пролиферативных процессов в эндометрии связана с увеличением количества рецепторов прогестерона на мембране клеток эндометрия, что усиливает в нем пролиферативные процессы под влиянием этого гормона. Наконец, увеличение концентрации эстрогенов в плазме крови стимулирует сокращение гладких мышц и микроворсин фаллопиевых труб, что способствует продвижению сперматозоидов в направлении ампулярного отдела фаллопиевых труб, где должно произойти оплодотворение яйцеклетки.
16.3.2.3.Секреторная фаза
Секреторная фаза менструального цикла начинается непосредственно после овуляции и продолжается до начала менструации (в среднем 12—16 дней). Основная роль секреторной фазы состоит в подготовке слизистой оболочки матки к имплантации яйцеклетки. В эту фазу активно функционирует желтое тело, стероидогенные клетки которого секретируют этсрадиол-17р, эстрон и прогестерон. Основным условием нормального течения секреторной фазы является высокая концентрация прогестерона в плазме крови. Прогестерон совместно с эстрогенами действует на клетки желез слизистой оболочки матки, вызывая в них секрецию. В секрете желез повышается уровень гликогена, гликопротеинов, гликолипидов, которые необходимы для поддержания метаболизма оплодотворенной яйцеклетки и способствуют ее внедрению в слизистую оболочку стенки матки. Если не происходит оплодотворения яйцеклетки и маточный цикл не завершается беременностью, менструальное желтое тело подвергается лютеолизу. Это вызывает снижение секреции клетками желтого тела эстрогенов и прогестерона и, как следствие, уменьшение концентрации в плазме крови женщины содержания этих гормонов. Понижение концентрации эстрогенов и прогестерона в плазме крови вызывает в слизистой оболочке функционального слоя миометрия повышение тонуса гладкомышечных клеток спиральных артерий, их скручивание и уменьшение просвета. 0 результате ишемии эндометрия возникает его некроз с последующим менструальным кровотечением.
16.3.3. Женский половой акт
Женский половой акт, как и мужской, состоит из стадии возбуждения, эрекции и разрешения. По аналогии с мужчиной, у женщины способность к осуществлению полового акта зависит от психической сексуальной стимуляции и от стимуляции эрогенных зон.
Половое возбуждение у женщины возникает при эротических переживаниях, вызванных ментальными процессами или просматриванием эротических картин или фильмов. Степень сексуального возбуждения варьирует на протяжении овариального цикла; половое влечение выше накануне овуляции, когда в крови увеличивается концентрация эстрогенов. Половое возбуждение у женщины тесно связано с рецептивной направленностью женской сексуальности, зависящей, от ситуации общения и эротической стимуляции со стороны мужчины (словесные, а также тактильные ласки, особенно эрогенных зон). Стимуляция эрогенных зон у женщин, связанная с раздражением рецепторов наружных половых органов (половых губ, преддверия влагалища и, особенно, клитора), в том числе во время полового акта, активирует многочисленные механорецепторы, сигналы от которых по срамным нервам поступают в парасимпатические центры крестцовых сегментов спинного мозга. Возбуждение парасимпатических эфферентных сосудорасширяющих нервов, а также симпатических вазоактивных волокон, идущих от нейронов грудных и поясничных сегментов к сосудам наружных и внутренних половых органов женщины, вызывает их кровенаполнение.
Основными проявлениями стадии возбуждения являются эрекция клитора и секреция слизи. Строение клитора идентично строению ткани мужского полового члена. Поэтому активация иннервирующих клитор парасимпатических волокон вызывает расслабление гладких мышц артерий при участии оксида азота, расширение просвета артерий и увеличение кровенаполнения ткани клитора. Импульсы по парасимпатическим нервам достигают также бартолиниевых желез, локализованных в глубине малых половых губ, и вызывает секрецию слизи. Слизь, секретируемая бартолиниевы- ми железами и слизистой оболочкой влагалища, а также слизь, выделяющаяся из желез мужской уретры во влагалище во время полового акта, способствует увлажнению влагалища и формированию оптимального сексуального чувства во время движений пениса во влагалище. Во время полового акта усиление кровенаполнения органов малого таза по мере нарастания нервного возбуждения изменяет размеры и положение матки, которая отклоняется к задней стенке влагалища, образуя в области шейки матки так называемую семяпринимающую полость. Одновременно в нижней части влагалища в результате застоя крови может формироваться оргастическая манжетка.
Оргазм. У женщин ощущение оргазма формируется в результате рефлекторных сокращений внутренних (например, ритмические сокращения оргастической манжетки влагалища под влиянием симпатических рефлекторных механизмов) либо наружных половых органов (клитор). При этом интенсивность мышечных сокращений во время оргазма прямо взаимосвязана с увеличением концентрации в плазме крови окситоцина. Секреция окситоцина из клеток нейрогипофиза во время оргазма регулируется миндалевидным комплексом и гипоталамусом. Ритмические сокращения мышц промежности у женщины во время оргазма регулируются также рефлекторно центрами спинного мозга крестцовых сегментов. У мужчин нейроны этих же крестцовых сегментов рефлекторно регулируют эякуляцию. Во время оргазма у женщины рефлекторные сокращения гладких мышц внутренних половых органов, а также расслабление шейки матки способствуют продвижению спермы через матку и фаллопиевы трубы.
Интенсивное сексуальное чувство, которое возникает у женщины во время оргазма, сопряжено с активацией двигательных систем головного мозга, что сопровождается повышением тонуса скелетных мышц. Во время оргазма увеличиваются частота дыхания, системное артериальное давление и частота сердцебиений. Стадия оргазма у женщины, так же как и у мужчин, сменяется стадией разрешения, во время которой нормализуются параметры систем кровообращения и дыхания. Однако у женщин отсутствует период рефрактерности после оргазма.








