 2020-06-08
2020-06-08 336
336Основную массу поджелудочной железы составляют ее экзокринные элементы, 80—95 % которых приходится на ацинозные (ацинарные) клетки, секретирующие ферменты. Центроацинозные и протоковые клетки секретируют воду, электролиты, слизь; из протоков компоненты смешанного секрета частично реабсорбируются.
Поджелудочная железа человека натощак выделяет небольшое количество-панкреатического секрета, (0»2—0,3.мл/мин), а после приема пищи 4—4,5 мл/мин. За сутки выделяется 1,5—2,5 л бесцветного прозрачного сока сложного состава.
Среднее содержание воды в соке 987 г/л. Основность сока (pH 7,5— 8,8) ооусловлена гидрокарбонатом (25—150 ммоль/л), концентрация которого в соке изменяется прямо пропорционально скорости секреции. В соке содержатся хлориды (4—130 ммоль/л) натрия и калия; между концентрацией гидрокарбонатов и хлоридов существует обратная зависимость, что связано с механизмом образования гидрокарбонатов клетками протока железы (рис. 8.13). Гидрокарбонаты панкреатического секрета участвуют в нейтрализации кислого пищевого содержимого желудка в двенадцатиперстной кишке. Соли кальция составляют 1—2,5 ммоль /л, В соке значительна концентрация белка (2—3,5 г/л), основную часть которого составляют ферменты., переваривающие все виды питательных веществ (табл. 8.4).
Та б л и ц а 8.4. Ферменты сока поджелудочной железы человека
| Протеолитические: | Амилолитические: |
| Трипсин(оген) I, II, Ш Химотрипсин(оген) А, В, С (Про)карбоксипептидаза Аь А3 (Про)карбоксипептидаза Bh В2 (Про)эластаза 1, 2 Липолитические: Липаза (Про)фосфолипаза Аь А2 | Неспецифическая эстераза | а-Амилаза Нуклеазы: Рибонуклеаза Дезоксирибонуклеаза Другие ферменты: Колипаза 1,2 Ингибитор трипсина Щелочная фосфатаза ___________________________________ |
Амилаза, липаза, колипаза, щелочная фосфатаза, ингибитор трипсина и нуклеазы секретируются поджелудочной железой в активном состоянии, а протеазы и фосфолипазы — в виде зимогенов.
|
|

 |
Трипсиноген поджелудочного сока в двенадцатиперстной кишке под действием ее фермента энтерокиназы (энтеропептидаза) превращается в трипсин, который вызывает последующую активацию трипсиногена. Активация состоит в отщеплении от трипсиногена гексапептида под действием энтерокиназы и трипсина при pH 6,8—8,0. Процесс ускоряется в присутствии Са2+
Химотрипсиноген, прокарбоксипептидазы А и В, проэластазы и профосфолипазы активируются трипсином. Трипсин, химотрипсин и эластаза расщепляют преимущественно внутренние пептидные связи белков и высокомолекулярных полипептидов. В результате этого образуются в основном низкомолекулярные пептиды.
Сок поджелудочной железы содержит а-амилазу, расщепляющую полисахариды. Производные нуклеиновых кислот расщепляют рибо- и дезоксирибонуклеазы. Липаза расщепляет жиры, в основном триглицериды, до моноглицеридов и жирных кислот. На липиды действуют также фосфолипаза А2 и эстераза.
Поскольку триглицериды нерастворимы в воде, липаза действует на поверхности жира. Чем больше поверхность жира, тем активнее идет его гидролиз. Поэтому эмульгирование жира имеет огромное значение для его переваривания. Эмульгирование обеспечивается желчью. Активность липазы повышает фермент колипаза. Она связывается с липазой в присутствии желчных солей и снижает оптимум pH действия фермента с 9 до 6—7, а также способствует адсорбции липазы на слизистой оболочке кишки.
Регуляция секреции поджелудочной железы. Секреция поджелудочной железы регулируется нервными и гуморальными механизмами. Раздражение блуждающих нервов вызывает выделение поджелудочного сока, богатого ферментами. Холинергические волокна блуждающих нервов посредством ацетилхолина действуют на М|-холинорецепторы панкреацитов и стимулируют секрецию ими ферментов и гидрокарбонатов. Холинергические
нейроны также потенцируют секреторные эффекты секретина и холецистокинина. Хирургическая ваготомия существенно снижает секрецию поджелудочной железы.
Симпатические волокна, иннервирующие поджелудочную железу, через p-адренорецепторы тормозят поджелудочную секрецию, усиливают синтез органических веществ в ней. Эффекты снижения секреции обеспечиваются также уменьшением кровоснабжения поджелудочной железы путем сужения кровеносных сосудов через их а-адренорецепторы.
Торможение секреции вызывают болевые раздражения, сон, напряженная физическая и умственная работа и др.
Поджелудочная железа имеет также пептидергическую иннервацию-, окончания этих нейронов выделяют ряд нейропептидов. Одни из них стимулируют (GRP, VIP, PHI и др.), а другие тормозят (энкефалин, PYY и др.) секрецию.
Гуморальная стимуляция панкреатической секреции. Первым открытым и названным гормоном веществом явился стимулятор обиль
ного по объему сокоотделения и секреции гидрокарбонатов. Высвобождение этого гормона в кровь дуоденальными S-клетками происходит при действии на слизистую двенадцатиперстной кишки перешедшего в нее кислого желудочного содержимого. Секретин в большей мере стимулирует через соответствующие мембранные рецепторы и вторичные мессенджеры (аденилатциклаза и цАМФ) центроацинозные и протоковые клетки, в меньшей мере — ацинарные клетки, поэтому выделяется секрет с высокой концентрацией в нем гидрокарбонатов и низкой ферментативной активностью.
Секрецию гидрокарбонатов и воды усиливаю т также ВИП, нейротензин, гастринрилизинг-пептид; тормозят вещество П, соматостатин, простагландины (Е), пептид YY, кальцитонин, глюкагон, ПП.
Гормоном, усиливающим секрецию поджелудочной железы, является холеци стокинин (XIIK). Высвобождение гормона в кровь из ССК-клеток слизистой оболочки двенадцатиперстной и тощей кишки происходит под влиянием химуса, особенно продуктов начального гидролиза пищевого белка и жира, углеводов, некоторых аминокислот. Стимулируют высвобождение ХЦК Са2+ и снижение pH содержимого двенадцатиперстной кишки.
ХЦК действует преимущественно на ациноциты поджелудочной железы, поэтому выделяющийся в ответ на этот гормон сок богат ферментами. Одновременное действие на железу секретина и ХЦК, имеющее место при приеме пищи, усиливает друг друга. Пептид химоденин стимулирует секрецию химотрипсиногена. Секрецию ациноцитов усиливают также гастринрилизинг-пептид и гастрин, секретин, инсулин, NO, ВИП и некоторые менее выраженные стимуляторы секреции ферментов. Торм озят глюкагон ^со- матостатин, вещество П^ э нке фалин, ГИП, ПП, пептид YY, кальцИТбнин.
~АциТюциты имеют на базолатеральнои мембране-несколько типов специфических рецепторов, на которые действуют нейротрансмиттеры и гормоны. В зависимости от их видов включаются две цепи вторичных посредников (рис. 8.14). В первой их цепи, мобилизуемой секретином и ВИП через их рецепторы, последовательно включаются аденилатциклаза (АЦ), цАМФ и соответствующая протеинкиназа, посредством которой активируется экзоцитоз гранул зимогена. Во второй цепи воздействие на мембранные рецепторы ацетилхолина (АЦХ), ХЦК и др. мобилизует фосфатидилинозитолбифосфат (ФИФг), а он в свою очередь — инозитол 1,4,5-три- фосфат (ИФЗ) и 1,2-диацилглицерол. Под их действием из ретикулума вы-
Секреция
пепсиногена
|
|

 |
свобождаются Са2+, в их присутствии оксидазотсинтаза (NOS) выделяет из L-аргинина оксид азота (N0), который активирует цГМФ, а последний активирует специфическую для него протеинкиназу, которая и обеспечивает экзоцитоз гранул зимогена через апикальную мембрану ациноцита.
Фазы секреции поджелудочной железы. Секреция поджелудочного сока резко усиливается через 2—3 мин после приема пищи и продолжается 6—14 ч. От количества и качества пищи зависят объем выделяющегося сока, его состав и динамика. Чем выше кислотность пищевого содержимого желудка, поступающего в двенадцатиперстную кишку, тем больше выделяется поджелудочного сока и гидрокарбонатов в его составе. Поэтому динамика поджелудочной и желудочной секреции схожи.
Фазы панкреатической секреции при стимуляции ее приемом пищи те же, что для желудочной секреции. Но в отличие от них более выражены гормональные влияния на поджелудочную железу, особенно в кишечную фазу.
Первая, мозговая, фаза секреции поджелудочной железы вызывается видом, 'за пахом пищи и другими раздражителями, связанными с приемом пищи (условнорефлекторные раздражения), а также воздействиями на рецепторы слизистой оболочки рта, жеванием и глотанием (безусловнорефлекторные раздражения). У человека с фистулой поджелудочного протока наблюдали условнорефлекторное выделение панкреатического сока через 2—3 мин после того, как испытуемому говорили о пище, которую ему дадут. Нервные импульсы, возникающие в рецепторах, достигают продолговатого мозга и затем по волокнам блуждающего нерва поступают к железе и вызывают ее секрецию.
По объему секреции первая фаза составляет в среднем 15 % общей по- стпрандиальной (послетрапезной) секреции поджелудочной железы, а по секреции ферментов — 25 %. Это подтверждает большую роль рефлекторного вагусного механизма в стимуляции ферментовыделительной деятельности железы. Секреция ее в первую фазу вариабельна и в большой мере зависит от аппетита, вида принимаемой пищи.
Вторая Фаза — желудочная. Секреция во время нее стимулируется и поддерживается путем ваго-вагального рефлекса с механо- и хеморецепторов желудка и посредством гастрина. На нее приходится около 10 % общей секреции.
С переходом желудочного содержимого в двенадцатиперстную кишку начинается третья — кишечная фаза панкреатической секреции. Она составляет 70—80 % общей секреции. В эту фазу секреция стимулируется посредством ваго-вагального дуоденопанкреатического рефлекса, секретина и ХЦК. Высвобождение секретина и ХЦК из продуцирующих их клеток происходит при действии на слизистую оболочку двенадцатиперстной кишки ее кислого содержимого и продуктов гидролиза нутриентов. Секреция ферментов в кишечную фазу стимулируется посредством ваго-вагального рефлекса и ХЦК. ХЦК и другие пептиды поддерживают этот рефлекс, стимулируя и потенцируя рецепторы вагусных афферентов. Кишечная фаза слагается из дуоденальной (основной), подвздошно-ободочной (особенно выражена, если этого отдела кишечника достигают недостаточно гидролизованные нутриенты, которые тормозят не только секрецию железы, но и скорость продвижения химуса по тонкой кишке) и циркуляторной (вызывается всосавшимися в кровь продуктами гидролиза нутриентов, выступающими в роли стимуляторов железы и потребляемых ею энергетических веществ).
В кишечную фазу велика роль саморегуляции панкреатической секреции, которая зависит от ферментных свойств дуоденального содержимого. «Избыток» в нем ферментов селективно по принципу отрицательной обратной связи тормозит секрецию ферментов. Избыток субстрата снимает эти тормозные влияния. Образовавшиеся продукты гидролиза субстрата стимулируют секрецию соответствующих ферментов поджелудочной железой. Эти механизмы направлены на срочную адаптацию секреции панкреатических ферментов к виду принятой пищи. Их реализация обеспечивается М-холинергическими и p-адренергическими влияниями, ХЦК и секретином. При стимуляции секреции поджелудочной железы усиливается ее кровоснабжение, что важно для поддержания секреции железы на высоком уровне.
Влияние пищевых режимов на секрецию поджелудочной железы. Прием пищи вызывает увеличение выделения всех ферментов в составе сока, но при углеводной пище в наибольшей мере увеличивается секреция амилазы, белковой пищи — трипсина и химотрипсина, прием жирной пищи вы
зывает секрецию сока с более высокой липолитической активностью. Набор ферментов в панкреатическом соке срочно адаптируется к составу принимаемой пищи во все три фазы секреции, и особенно в ее кишечную фазу.
Есть медленные адаптации секреции ферментов под влиянием длительного приема пищи определенного состава. Суть адаптации состоит в том, что железа секретирует больше того фермента, который гидролизует преобладающие в рационе питательные вещества. Эта адаптация носит согласованный характер с работой других отделов пищеварительного тракта, составляя часть интегрированных адаптаций всего пищеварительного конвейера.
8.8.2. Желчеобразование и желчевыделение
Участие желчи в пищеварении. Желчь образуется в печени; ее участие в пищеварении многообразно. Желчь эмульгирует жиры, увеличивая поверхность, на которой осуществляется их гидролиз липазой; растворяет продукты гидролиза жиров, способствует их всасыванию и ресинтезу триглицеридов в энтероцитах; повышает активность панкреатических и кишечных ферментов, особенно липазы. Желчь усиливает гидролиз и всасывание белков и углеводов, всасывание жирорастворимых витаминов, холестерина и солей кальция; является стимулятором желчеобразования, желчевыделения, моторной и секреторной деятельности тонкой кишки, апоптоза и пролиферации энтероцитов.
Состав желчи и ее образование. У человека за сутки образуется около 1—2 л желчи. Процесс образования желчи — желчеотделение (холерез) — идет непрерывно, а поступление желчи в двенадцатиперстную кишку — желчевыделение (холекинез) — периодически, в основном в связи с приемом пищи. Натощак желчь в кишечник почти не поступает, а направляется в желчный пузырь, где при депонировании концентрируется и изменяет свой состав. Поэтому принято говорить о двух видах желчи — печеночной и пузырной.
Желчь является не только секретом, но и экскретом. В ее составе выводятся различные эндогенные и экзогенные вещества (табл. 8.5). В желчи содержатся белки, аминокислоты, витамины и другие вещества. Желчь обладает небольшой ферментативной активностью, pH печеночной желчи 7,3—8,0. При прохождении желчи по желчевыводящим путям и нахождении в желчном пузыре жидкая и прозрачная золотисто-желтого цвета печеночная желчь с относительной плотностью 1,008—1,015 концентрируется, так как из нее всасываются вода и минеральные соли, к ней добавляется муцин желчных путей и пузыря, и желчь становиться темной, тягучей, увеличивается ее относительная плотность до 1,026—1,048 и снижается pH до 6,0—7,0 за счет образования солей желчных кислот и всасывания гидрокарбонатов. Основное количество желчных кислот и их солей содержится в желчи в виде соединений с гликоколом и таурином.
Желчные пигменты являются продуктами распада гемоглобина и других производных порфиринов. Основным желчным пигментом человека является билирубин — пигмент красно-желтого цвета, придающий печеночной желчи характерную окраску. Другой пигмент зеленого цвета — биливердин в желчи человека содержится в следовых количествах.
Желчь образуется гепатоцитами (примерно 75 % ее объема) и эпителиальными клетками желчных протоков (около 25 % ее объема).
Таблица 8.5. Состав желчи
| Компоненты | Печеночная желчь | Пузырная желчь |
| Вода (г/л) | 950-980 | 850-920 |
| Сухой остаток (г/л): Соли желчных кислот | 10,0-11,0 | 30-100 |
| Жирные кислоты | 1,0 | 3,0-12,0 |
| Билирубин | 2,0 | 5,0-20,0 |
| Холестерин | 0,5-1,0 | 3,0-9,0 |
| Лецитин | 0,4 | 1,0-4,0 |
| Неорганические соли | 8,0-8,5 | 6,0-6,5 |
| Ионы (ммоль/л): Na+ | 145 | 130 |
| К+ | 5 | 9 |
| Са2+ | 2,5 | 6 |
| er | 100 | 75 |
| HCOJ | 1 28 | 10 |
Желчные кислоты синтезируются в гепатоцитах. Из тонкой кишки всасывается в кровь около 85—90 % желчных кислот, выделившихся в кишку в составе желчи. Всосавшиеся желчные кислоты с кровью по воротной вене приносятся в печень и включаются в состав желчи (энтеропанкреати- ческая циркуляция). Остальные 10—15 % желчных кислот выводятся из организма в основном в составе кала. Эта потеря желчных кислот восполняется их синтезом в гепатоцитах.
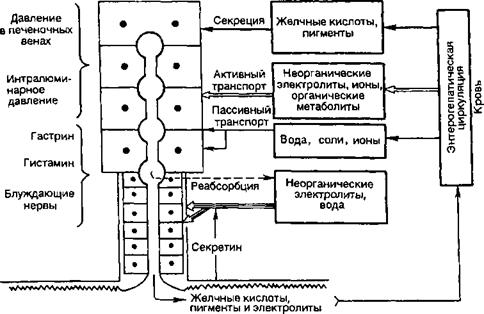
В целом образование желчи происходит путем активной секреции компонентов желчи (желчные кислоты) гепатоцитами, активного и пассивного транспорта веществ из крови через клетки и межклеточные контакты (вода, глюкоза, креатинин, электролиты, витамины, гормоны и др.) и обратного всасывания воды и ряда веществ из желчных капилляров, протоков и желчного пузыря (рис. 8.15). Ведущая роль в образовании желчи принадлежит секреции.
Регуляция желчеобразования. Желчеобразование идет непрерывно, но его рефлекторно и гуморально усиливают акт еды и принятая пища. Парасимпатические холинергические влияния усиливают, а симпатические адренергические снижают желчеобразование. К числу гуморальных стимуляторов желчеобразования (холеретики) относится сама желчь. Секретин усиливает секрецию желчи, выделение в ее составе воды и электролитов (гидрокарбонаты). Слабее стимулируют желчеобразование глюкагон, гастрин и ХЦК.
Желчевыделение. Движение желчи в желчевыделительном аппарате обусловлено разностью давления в его частях и двенадцатиперстной кишке, состоянием сфинктеров внепеченочных желчных путей. Выделяют 3 сфинктера: в месте слияния пузырного и общего печеночного протока (Мириззи), в шейке желчного пузыря (Люткенса) и концевом отделе общего желчного протока (Одди). Тонус мышц этих сфинктеров определяет направление движения желчи. Давление в желчевыделительном аппарате создается секреторным давлением желчеобразования и сокращениями гладких мышц протоков и желчного пузыря. Эти сокращения согласованы
Тонкая кишка
Рис. 8.15. Желчеобразование и его регуляция.
с тонусом сфинктеров и регулируются нервными и гуморальными механизмами. Давление в общем желчном протоке колеблется от 4 до 300 см вод.ст. В желчном пузыре давление вне пищеварения составляет 60—185 см вод.ст.; во время пищеварения за счет сокращения пузыря оно поднимается до 200—300 см вод.ст., обеспечивая выход желчи в двенадцатиперстную кишку через открывшийся сфинктер Одди.
Вид, запах пищи, подготовка к ее приему и сам прием вызывают сложные изменения деятельности желчевыделительного аппарата. Желчный пузырь при этом через различный латентный период сначала расслабляется, а затем сокращается, и желчь в небольшом количестве выходит в двенадцатиперстную кишку. Этот период первичной реакции желчевыделительного аппарата длится 7—10 мин. На смену ему приходит основной эва- куаторный период, во время которого сокращение желчного пузыря чередуется с расслаблением и через открытый сфинктер Одди переходит в двенадцатиперстную кишку сначала желчь из общего протока, затем пузырная, а в последующем — печеночная желчь. Сильными возбудителями желчевыделения являются яичный желток, молоко, мясо и жиры.
Рефлекторная стимуляция желчевыделительного аппарата и холекинеза осуществляется условно- и безусловнорефлекторно через блуждающие нервы при раздражении рецепторов ротовой полости, желудка и двенадцатиперстной кишки.
Большую роль в стимуляции желчевыделения играет ХЦК, вызывающий сокращения желчного пузыря. Слабые сокращения его вызывают гастрин, секретин, ГРП. Тормозят сокращения желчного пузыря глюкагон, кальцитонин, ВИП, ПП, антихолецистокинин.
8.8.3. Кишечная секреция
Кишечный сок представляет собой мутную вязкую жидкость и является продуктом деятельности всей слизистой оболочки тонкой кишки; имеет сложный состав и разное происхождение. За сутки у человека выделяется до 2,5 л сока тонкой кишки.
В криптах слизистой оболочки двенадцатиперстной кишки заложены брушерсеы^железы. Их сок — густая бесцветная жидкость слабооснбвной реакции; обладает небольшой протеолитической,-амилолитической и липолитической активностью. Либеркюновы железы, или кишечные крипты, заложены в слизистой оболочке двенадцатиперстной и всей тонкой кишки. Секреторной способностью обладают многие эпителиоциты крипт тонкой кишки.
Эпителиоциты с исчерченной каемкой покрывают ворсинку. Апикальная часть клетки содержит многие кишечные ферменты, которые трансло- цированы из клетки, где они были синтезированы. Ферментами богаты также лизосомы.
Бокаловидные кде/пк/д-выделяют слизь через разрывы апикальной плазматической мембраны. Секрет обладает ферментативной активностью.
Энтероциты с ацидофильными гранулами, кле тки Панета, в зрелом состоянии также имеют морфологические признаки мерокриновой и апокриновой секреции. Секрет содержит гидролитические ферменты. В криптах заложены также аргентаффинные клетки, выполняющие эндокринные функции.
Находящееся в полости петли тонкой кишки содержимое является продуктом многих клеток и процессов, в том числе десквамации энтероцитов и двустороннего транспорта высоко- и низкомолекулярных веществ. Это, собственно, и есть то, что называется кишечным соком.
Свойства и состав кишечного сока. Центрифугирование кишечного сока разделяет его на жидкую и плотную части. Жидкая часть сока образована секретом, транспортированными из крови растворами неорганических и органических веществ и содержимым разрушенных ’ энтероцитов, имеет около 20 г/л сухого вещества. В числе неорганических веществ (10 г/л) — хлориды, гидрокарбонатьптфосфат ы натрия, калия, кальция. Сок имеет pH 7,2—7,5: при усилении секреции — до 8.6. Органические вещества представлены слизью, белками, аминокислотами, мочевиной и другими продуктами обмена веществ. Плотная часть сока — желтовато-серая масса, имеет вид слизистых комков и включает в себя неразрушенные эпителиальные клетки, их фрагменты и слизь — секрет бокаловидных клеток.
Слизь образует защитный слой, предотвращающий чрезмерное механическое и химическое воздействие химуса на слизистую оболочку кишки. В слизи высока активность пищеварительных ферментов.
В слизистой оболочке тонкой кишки идет непрерывная смена слоя клеток поверхностного эпителия. Они образуются в криптах, затем продвигаются по ворсинкам и слущиваются с их верхушек — морфокинетическая (или морфонекротическая) секреция. Полное обновление этих клеток у человека совершается в среднем за 3—5 сут. Высокий темп образования и отторжения клеток обеспечивает достаточно большое их количество в кишечном соке: за сутки отторгается около 250 г эпителиоцитов.
Плотная часть сока обладает значительно большей ферментативной активностью, чем жидкая. В кишечном соке содержится более 20 ферментов. Кишечные ферменты обеспечивают пристеночное пищеварение. Углеводы гидролизуются а-глюкозидазами, а-галактазидазой (лактаза), глюкоамила
зой (у-амилаза). К ct-глюкозидазам относятся мальтаза; дисахаридазы: изомальтаза, сахараза (инвертаза) и трегалаза. С их участием из соответствующих дисахаридов образуются моносахариды. Дефицит кишечных дисахаридаз приводит к непереносимости соответствующего дисахарида. Известны генетически закрепленные и приобретенные лактазная, трегалазная, сахаразная и комбинированные ферментные недостаточности.
В тонкой кишке продолжается и завершается гидролиз пептидов. Аминопептидазы составляют основную часть пептидазной активности щеточной каймы энтероцитов и расщепляют пептидную связь между двумя определенными аминокислотами, которые затем всасываются. В пристеночном гидролизе липидов важное значение имеет кишечная моноглице- ридлипаза. Она гидролизует моноглицериды с любой длиной углеводородной цепи, а также короткоцепочечные ди- и триглицериды, в меньшей мере — триглицериды со средней длиной цепи и эфиры холестерина. Начальный гидролиз нуклеопротеидов осуществляется протеазами, затем гидролизуются отщепленные от белковой части РНК и ДНК соответственно РНК- и ДНКазами до олигонуклеотидов, которые затем при участии нуклеаз и эстераз деградируют до нуклеотидов. Последние атакуются щелочными фосфатазами и более специфичными нуклеотидазами с высвобождением всасываемых затем нуклеозидов. Фосфатазная активность кишечного сока высока.
Ферментный спектр слизистой оболочки тонкой кишки и ее сока изменяется под влиянием длительных режимов питания.
Регуляция кишечной секреции. Секреция кишечных желез усиливается при приеме пищи, местном механическом и химическом раздражении кишки, посредством холинергических нейронов и под влиянием некоторых кишечных гормонов.
Механическое раздражение слизистой оболочки тонкой кишки увеличивает выделение жидкой части сока. Химическими стимуляторами секреции тонкой кишки являются продукты переваривания белка, жира, панкреатический сок, соляная и другие кислоты. Местное воздействие продуктов переваривания питательных веществ вызывает отделение кишечного сока, богатого ферментами.
Акт еды существенного влияния на кишечную секрецию не оказывает. Имеются данные о модуляции секреции под влиянием ЦНС, о стимулирующем действии на секрецию холиномиметических веществ, тормозном влиянии холинолитиков и симпатомиметических веществ. Стимулируют кишечную секрецию ГИП, ВИП, мотилин; тормозит — соматостатин.
8.8.4. Полостной и пристеночный гидролиз питательных веществ в тонкой кишке
В тонкой кишке происходит полостное и пристеночное пищеварение; не исключено и внутриклеточное.
Полостное пищеварение в тонкой кишке осуществляется ферментами панкреатического и кишечного секретов. В результате полостного пищеварения гидролизуются крупномолекулярные нутриенты и образуются в основном олигомеры. Последующий их гидролиз происходит по типу пристеночного пищеварения и завершается на мембране энтероцитов.
Регуляция полостного пищеварения осуществляется путем изменения секреции пищеварительных желез, скорости продвижения химуса по тонкой кишке, интенсивности пристеночного пищеварения и всасывания.
Регуляция пристеночного пищеварения изучена недостаточно. Интенсивность его зависит от полостного пищеварения и, следовательно, факторов, влияющих на него. Влияют на мембранное пищеварение гормоны надпочечников (синтез и транслокация ферментов), диеты и другие факторы. Пристеночное пищеварение зависит также от моторики кишки, изменяющей переход веществ из химуса в исчерченную каемку, величины пор исчерченной каемки, ферментного состава в ней, сорбционных свойств мембраны.
8.8.5. Моторная деятельность тонкой кишки
Моторная деятельность тонкой кишки обеспечивает перемешивание ее химуса с пищеварительными секретами, продвижение его по кишке, смену его у слизистой оболочки, повышение внутрикишечного давления, т.е. способствует гидролизу и всасыванию питательных веществ.
Движение тонкой кишки происходит в результате координированных сокращений продольного и циркулярного слоев гладких мышц. Принято различать несколько типов сокращений тонкой кишки (рис. 8.16): ритмическая сегментация, маятникообразные, перистальтические (очень медленные, медленные, быстрые, стремительные), антиперистальтические и тонические. Первые два типа относятся к ритмическим, или сегментирующим, сокращениям.
Ритмическая сегментация обеспечивается преимущественно сокращениями циркулярного слоя мышц, при этом содержимое кишки делится на части. Следующим сокращением образуется новый сегмент кишки, содержимое которого состоит из двух частей соседних сегментов. Данными сокращениями достигается перемешивание химуса.
Маятникообразные сокращения обеспечиваются продольными и циркулярными мышцами. При этом происходит перемещение химуса «вперед-назад» и слабое поступательное движение его в аборальном направлении. В верхних отделах тонкой кишки человека частота ритмических сокращений составляет 9—12, в нижних — 6—8 в 1 мин.
Перистальтическая волна, состоящая из перехвата и расширения тонкой кишки, продвигает химус в аборальном направлении. Одновременно по длине кишки продвигается несколько волн со скоростью 0,1—0,3 см/с, в проксимальных отделах быстрее, чем в дистальных. Скорость стремительной пропульсивной волны 7—12 см/с.
При антиперистальтических сокращениях волна движется в обратном, оральном направлении. В норме тонкая кишка, как и желудок, антиперистальтически не сокращаются (это характерно для рвоты).
Тонические сокращения могут иметь локальный характер или передвигаться с очень малой скоростью. Исходное (базальное) давление в полости тонкой кишки составляет 5—14 см вод.ст. Монофазные волны повышают внутрикишечное давление до 30—90 см вод.ст. Медленный компонент сокращений длится от одной до нескольких минут и повышает давление не столь значительно.
Моторика тонкой кишки регулируется миогенными, нервными и гуморальными механизмами. Миогенные механизмы обеспечивают автоматизм кишечных мышц и сократительную реакцию на растяжение кишки. Фазная сократительная деятельность кишки реализуется нейронами миэнтерального нервного сплетения, обладающими ритмической фоновой активностью. Кроме осцилляторов энтеральных метасимпатических

Рис. 8.16. Типы сокращений тонкой кишки.
а перистальтика, б — сегментация. Стрелки — направления движения химуса.
ганглиев имеются два датчика ритма кишечных сокращений — первый у места впадения в двенадцатиперстную кишку общего желчного протока, второй — в подвздошной кишке. Эти датчики и ганглии энтерального нервного сплетения контролируются нервными и гуморальными механизмами.
Парасимпатические влияния преимущественно усиливают, симпатические тормозят моторику тонкой кишки. Описаны пептидергические нервные влияния обоих типов. Эффекты раздражения вегетативных нервов в большой мере зависят от состояния кишки, на фоне которого производятся раздражения. Изменяют моторику раздражения спинного и продолговатого мозга, гипоталамуса, лимбической системы, коры большого мозга. Раздражения передних и средних ядер отделов гипоталамуса преимущественно возбуждают, а заднего тормозят моторику желудка, тонкой и толстой кишки.
Акт еды тормозит, а затем усиливает кишечную моторику. В дальнейшем она зависит от физических и химических свойств химуса: усиливают ее грубые виды пиши, богатые непереваренными в тонкой кишке пищевыми волокнами, продукты переваривания питательных веществ, особенно жиры, кислоты, основания, соли.
Важное значение имеют рефлексы с различных отделов пищеварительного тракта на моторику тонкой кишки: пищеводно-кишечный (возбуждающий), желудочно-кишечный (возбуждающий и тормозящий), ректоэнте- ральный (тормозящий). Дуги этих рефлексов замыкаются на различных уровнях ЦНС и в периферических ганглиях. В целом моторная деятельность любого участка тонкой кишки — суммарный результат местных, удаленных влияний в пределах пищеварительного тракта и влияний с других систем организма.
Моторику тонкой кишки усиливают, действуя на миоциты или энтеральные нейроны, серотонин, гистамин, гастрин, мотилин, ХЦК, вещество П, вазопрессин, окситоцин, брадикинин и др., тормозят — секретин, ВИП, ГИП и др.
8.8.6. Всасывание различных веществ
в тонкой кишке
Различные вещества всасываются в пищеварительном тракте посредством различных механизмов, имея характерную топографию всасывания.
Всасывание воды и минеральных солей. В желудочно-кишечный тракт в составе пищи и выпиваемых жидкостей поступает за 1 сут 2—2,5 л воды, в 434 составе секретов пищеварительных желез 6—7 л, выводится же с калом всего 100—150 мл воды. Остальное количество воды всасывается из пищеварительного тракта в кровь, небольшое количество — в лимфу. Всасывание воды начинается в желудке, наиболее интенсивно происходит в тонкой и особенно толстой кишке.
Основное количество воды всасывается из изотонических растворов кишечного химуса, так как в кишечнике гипер- и гипотонические растворы соответственно концентрируются или разводятся. Абсорбция воды из изотонических и гипертонических растворов требует затраты энергии. Всасываемые эпителиоцитами растворенные вещества «тянут» за собой воду. Решающая роль в переносе воды принадлежит ионам и особенно натрия. Поэтому все факторы, влияющие на его транспорт, изменяют и всасывание воды. Оно сопряжено также с транспортом сахаров и аминокислот. Поэтому многие эффекты замедления или ускорения всасывания воды являются результатом изменения транспорта из тонкой кишки других веществ.
Интенсивность всасывания натрия и воды в кишке максимальна при pH 6,8 (при pH 3,0 всасывание воды прекращается). Изменяют всасывание воды рационы питания. Увеличение в них доли белка повышает скорость всасывания воды, Na+ и С1_ Изменяется скорость всасывания воды в зависимости от гидратированности организма.
Доказано условнорефлекторное изменение всасывания воды; замедление под влиянием наркоза и после ваготомии, что свидетельствует о роли ЦНС в этом процессе. Влияют на всасывание воды многие гормоны желез внутренней секреции и некоторые гастроинтестинальные гормоны — снижают ее всасывание гастрин, секретин, ХЦК, ВИП, ГРП, серотонин.
За сутки в желудочно-кишечном тракте всасывается более 1 моля хлорида натрия. В желудке натрий почти не всасывается, но интенсивно всасывается в толстой и подвздошной кишке, в тощей кишке его всасывание значительно меньше.
Ионы Na+ поступают из полости тонкой кишки в кровь через кишечные эпителиоциты и между ними. Поступление Na+ в эпителиоцит происходит по электрохимическому градиенту пассивным путем. Имеется также система транспорта Na+, сопряженная с транспортом сахаров и аминокислот, возможно, и с О" и НСОз Ионы Na+ из эпителиоцитов через их базолатеральные мембраны транспортируются активно в межклеточную жидкость, кровь и лимфу. Это обеспечивает возможность дальнейшего пассивного транспорта Na+ через апикальные мембраны в эпителиоциты из полости кишечника. Различные стимуляторы и ингибиторы всасывания Na+ действуют прежде всего на механизмы активного транспорта базолатеральных мембран эпителиоцитов. Транспорт Na+ по межклеточным каналам совершается пассивно по градиенту концентрации. Интенсивность всасывания натрия зависит от pH кишечного содержимого, гидратации организма и содержания в нем этого элемента. Усиливают всасывание натрия минералокортикоиды (альдостерон), угнетают — гастрин, секретин и холецистокинин.
Всасывание калия происходит в основном в тонкой кишке посредством механизмов активного и пассивного транспорта по электрохимическому градиенту. Активный транспорт К+ сопряжен с транспортом Na+ в базолатеральных мембранах эпителиоцитов.
Всасывание хлора происходит в желудке и наиболее активно в подвздошной кишке по типу активного и пассивного транспорта. Пассивный транспорт С1“ сопряжен с транспортом Na+. Активный транспорт С1“ че
рез апикальные мембраны сопряжен с транспортом Na+ или обменом С1 на НСОз
Двухвалентные ионы в желудочно-кишечном тракте всасываются очень медленно. Так, в кишечник человека поступает ежесуточно 35 ммоль кальция, но только половина его всасывается. Кальций всасывается в 50 раз медленнее, чем Na+, но быстрее, чем двухвалентные ионы железа, цинка и марганца. Всасывание кальция совершается с участием переносчиков, активируется желчными кислотами и витамином D, поджелудочным соком, некоторыми аминокислотами, натрием, угнетается многими веществами. При недостатке кальция в организме его всасывание увеличивается, в чем большую роль могут играть гормоны ряда желез, но особенно паратирин.
Всасывание продуктов гидролиза белков. Белки всасываются в основном в кишечнике после их гидролиза до аминокислот. Всасывание различных аминокислот происходит с неодинаковой скоростью в различных отделах тонкой кишки. Быстрее других всасываются аргинин, метионин, лейцин; медленнее — фенилаланин, цистеин, тирозин и еще медленнее — аланин, серин, глутаминовая кислота. L-формы аминокислот всасываются интенсивнее, чем D-формы. Всасывание аминокислот из кишки через апикальные мембраны в эпителиоциты осуществляется активно посредством транспортеров со значительной затратой энергии фосфоросодержащих макроэргов. Количество всасывающихся пассивно аминокислот невелико.
Существует несколько видов транспортеров аминокислот в апикальных мембранах эпителиоцитов. Из эпителиоцитов аминокислоты транспортируются в межклеточную жидкость по механизму облегченной диффузии. Транспорт аминокислот через апикальную и базальную мембраны взаимосвязан. Большинство аминокислот, образующихся в процессе гидролиза белков и пептидов, всасывается быстрее, чем свободные аминокислоты, введенные в тонкую кишку. Транспорт натрия стимулирует всасывание аминокислот. Из менее концентрированных растворов аминокислот они всасываются быстрее, чем из более концентрированных.
Интенсивность всасывания аминокислот зависит от возраста, уровня белкового обмена в организме, содержания в крови свободных аминокислот и ряда других факторов, от нервных и гуморальных влияний.
Три- и дипептиды в тонкой кишке всасываются посредством специального транспортера апикальной мембраны.
Всасывание углеводов. Происходит в основном в тонкой кишке. С наибольшей скоростью всасываются гексозы; в их числе глюкоза и галактоза; пентозы всасываются медленнее. Всасывание глюкозы и галактозы использует механизм активного транспорта через апикальные мембраны кишечных эпителиоцитов. Транспорт моносахаридов, образующихся при гидролизе олигосахаридов, осуществляется с большей скоростью, чем всасывание моносахаридов, введенных в просвет кишки. Всасывание глюкозы (и некоторых других моносахаридов) через апикальные мембраны кишечных эпителиоцитов активируется транспортом натрия.
Глюкоза аккумулируется в кишечных эпителиоцитах, и последующий транспорт ее из них через базолатеральные мембраны в межклеточную жидкость и кровь происходит по градиенту концентрации, а также с участием специальных транспортеров.
Всасывание фруктозы (и некоторых других моносахаридов) не зависит от транспорта Na+ и происходит активно. Не исключают возможности и пассивного транспорта фруктозы.
Всасывание углеводов тонкой кишкой усиливается некоторыми аминокислотами, резко тормозится ингибиторами тканевого дыхания. Всасывание различных моносахаридов в различных отделах тонкой кишки происходит с неодинаковой скоростью. Так, скорость всасывания глюкозы в тощей кишке в 3 раза выше, чем в подвздошной.
На всасывание сахаров влияют диета, многие факторы внешней среды, концентрация глюкозы в крови. Существует сложная нервная и гуморальная регуляция всасывания углеводов. Доказано изменение их всасывания под влиянием коры и подкорковых структур головного мозга, его ствола и спинного мозга.
Парасимпатические влияния усиливают, а симпатические — тормозят всасывание углеводов. Всасывание глюкозы усиливается гормонами надпочечников, гипофиза, щитовидной железы, а также серотонином и ацетилхолином.
Гистамин незначительно, а соматостатин существенно тормозят всасывание глюкозы.
Всасывание продуктов гидролиза жиров. Всасывание липидов наиболее активно происходит в двенадцатиперстной кишке и проксимальной части тощей кишки. Скорость всасывания различных жиров зависит от их эмульгирования и гидролиза. В результате действия в полости кишки панкреатической липазы из триглицеридов образуются диглицериды, затем моноглицериды и жирные кислоты, хорошо растворимые в растворах солей желчных кислот. Кишечная липаза в зоне исчерченной каемки эпителиоцитов завершает гидролиз липидов. Из моноглицеридов, жирных кислот с участием солей желчных кислот, фосфолипидов и холестерина образуются мельчайшие мицеллы (диаметр их около 100 нм), которые через апикальные мембраны переходят в кишечные эпителиоциты. Желчные кислоты мицелл остаются в полости кишки и всасываются в подвздошной кишке по механизму активного транспорта.
В кишечных эпителиоцитах происходит ресинтез триглицеридов. Из них, а также холестерина, фосфолипидов и глобулинов образуются хиломикроны — мельчайшие жировые частицы, заключенные в белковую оболочку. Хиломикроны покидают эпителиоциты через базолатеральные мембраны, переходят в соединительные пространства ворсинок, оттуда проникают в центральный лимфатический сосуд ворсинки, чему содействуют ее сокращения.
Основное количество жира всасывается в лимфу, поэтому через 3—4 ч после приема пищи лимфатические сосуды наполнены лимфой, напоминающей молоко и называемой млечным соком.
В нормальных условиях в кровь поступает небольшое количество всосавшегося в кишечнике жира, представленного триглицеридами жирных кислот с короткой углеводородной цепочкой. В кровеносные капилляры из эпителиоцитов и межклеточного пространства могут транспортироваться и растворимые в воде свободные жирные кислоты и глицерин. Для всасывания жиров с короткими и средними углеводородными цепями жирных кислот образование в эпителиоцитах хиломикронов не обязательно. Небольшое количество хиломикронов может поступать и в кровеносные сосуды ворсинок.
На скорость гидролиза и всасывания жира влияет ЦНС. Парасимпатический отдел вегетативной нервной системы ускоряет, а симпатический — замедляет всасывание жиров. Ускоряют их всасывание гормоны коры надпочечников, щитовидной железы и гипофиза, а также дуоденальные гормоны — секретин и ХЦК.
8.9. ФУНКЦИИ ТОЛСТОЙ КИШКИ
8.9.1. Поступление кишечного химуса в толстую кишку
За сутки у человека из тонкой в толстую кишку порционно переходит 0,5—4 л химуса через илеоцекальный сфинктер — баугиниеву заслонку. Сфинктер выполняет роль клапана. Его расслаблению и раскрытию илеоцекального прохода способствуют сокращения продольных мышц тонкой и толстой кишки. При наполнении слепой кишки и ее растяжении сфинктер плотно закрывается и содержимое толстой кишки в норме в тонкую кишку не возвращается.
Вне пищеварения сфинктер закрыт. После приема пищи через 1—4 мин каждые 0,5—1 мин он открывается и химус небольшими порциями (до 15 мл) поступает в толстую кишку. Раскрытие сфинктера происходит рефлекторно: перистальтическая волна тонкой кишки повышает давление в ней и расслабляет сфинктер и обычно — пилорический сфинктер (бисфинктерный рефлекс). Повышение давления в толстой кишке увеличивает тонус илеоцекального сфинктера и тормозит поступление в нее содержимого тонкой кишки.
8.9.2. Роль толстой кишки в пищеварении
Пища почти полностью переваривается и всасывается в тонкой кишке. Небольшое количество веществ пищи, в том числе клетчатка и пектин, в составе химуса подвергаются гидролизу в толстой кишке. Гидролиз осуществляется ферментами химуса и сока толстой кишки, а клетчатка и пектин — ферментами микроорганизмов.
Сок толстой кишки выделяется в небольшом количестве вне раздражения кишки. Местное механическое раздражение слизистой увеличивает секрецию в 8—10 раз. Сок состоит из жидкой и плотной частей, имеет основную реакцию (pH 8,5—9,0). Плотную часть сока составляют слизистые комки из отторгнутых кишечных эпителиоцитов и слизи, секретируемой бокаловидными клетками.
Основное количество ферментов содержится в плотной части сока, их активность значительно меньше, чем в тонкой кишке. В соке толстой кишки нет энтерокиназы и сахаразы, щелочной фосфатазы в 15—20 раз меньше, чем в соке тонкой кишки. В небольшом количестве в составе сока выделяются катепсины, пептидазы, липаза, амилаза и нуклеазы. В зависимости от осмотического и гидростатического давления кишечного содержимого интенсивно всасывается вода (до 4—6 л и более за 1 сут). Химус постепенно превращается в каловые массы (за сутки выводится 200—350 г сформированного кала). При растительной пище их больше, чем при смешанной или мясной. При пище, богатой неперевариваемыми волокнами (целлюлоза, гемицеллюлоза, пектин, лигнин), количество кала увеличивается не только за счет них, но и ускорения передвижения химуса и формируемого кала, что предотвращает запоры.
8.9.3. Моторная деятельность толстой кишки
Весь процесс пищеварения у взрослого человека длится 1 —3 сут, из них наибольшее время приходится на пребывание остатков пищи в толстой кишке. Ее моторика обеспечивает резервуарную функцию — накопление содержимого, всасывание из него ряда веществ, в основном воды, продвижение его, формирование каловых масс и их удаление (дефекация). У здорового человека контрастная масса через 3—3,5 ч после ее приема начинает поступать в толстую кишку. Она заполняется в течение 24 ч и полностью опорожняется за 48—72 ч.
Начальный отдел толстой кишки имеет в среднем 8 циклов сокращений в I мин, а сигмовидная кишка — до 17—18 циклов в 1 мин.
Толстая кишка осуществляет сокращения нескольких типов: малые и большие маятникообразные, перистальтические и антиперистальтические, пропульсивные. Первые 4 типа сокращений перемешивают содержимое кишки и повышают давление в ее полости, что способствует сгущению содержимого путем всасывания воды. Сильные пропульсивные сокращения возникают 3—4 раза в сутки и продвигают кишечное содержимое в аборальном направлении.
Толстая кишка получает парасимпатическую инервацию в составе блуждающих и тазовых нервов и усиливает моторику путем условных и безусловных рефлексов при раздражении пищевода, желудка и тонкой кишки. Симпатические нервы проходят в составе чревных нервов и тормозят моторику кишки. Ведущее значение в организации ее моторики имеют интрамуральные нервные механизмы при местном раздражении толстой кишки ее содержимым. Раздражение механорецепторов прямой кишки тормозит моторику вышележащих отделов тонкой кишки. Тормозят моторику толстой кишки серотонин, адреналин, глюкагон.
8.9.4. Газы толстой кишки
За сутки выводится из кишечника при дефекации и вне ее 100—150 мл газа. При метеоризме объем его может достигать Зли более. Газы кишечника имеют различное происхождение. Часть их попадает в пищеварительный тракт при заглатывании пищи, но в основном газы образуются в кишечнике. При взаимодействии гидрокарбонатов пищеварительных секретов с кислыми продуктами кишечного химуса образуется значительное количество СО2. Газы продуцирует и микрофлора кишечника. Некоторые виды пиши (бобовые, капуста, лук, черный хлеб, картофель и др.) и микрофлора способствуют образованию большого количества газов. У здоровых людей газовую смесь составляют азот (24—90 %), углекислый газ (4,3—29 %), кислород (0,1—23 %), водород (0,6 %), метан (0—26 %); в небольшом количестве сероводород, аммиак, меркаптан.
8.9.5. Дефекация
Дефекация — опорожнение толстой кишки от каловых масс, происходит в результате раздражения рецепторов прямой кишки накопившимися в ней каловыми массами. Позыв на дефекацию возникает при повышении давления в прямой кишки до 40—50 см вод.ст. Сфинктеры прямой кишки — внутренний, состоящий из гладких мышц, и наружный, из исчерченных мышц, вне дефекации находятся в состоянии тонического сокращения. В результате рефлекторного расслабления этих сфинктеров и перистальтических сокращений кал выводится из прямой кишки. В этом процессе большое значение имеет так называемое натуживание, при котором сокращаются мышцы брюшной стенки и диафрагмы, повышается внутри-

|
|
|
|
|
|
|
|
|
|

 Рис. 8.17. Рефлекторное обеспечение дефекации.
Рис. 8.17. Рефлекторное обеспечение дефекации.
Р — рецепторы; 1 — внутренний сфинктер заднего прохода; 2 — наружный сфинктер заднего прохода; 3 — анальные железы; 4 — интрамуральные ганглии; (+) — влияния, повышающие тонус мышцы; (—) — влияния, понижающие тонус мышцы.
брюшное давление, достигающее при акте дефекации 220 см вод.ст.; сокращается мышца, поднимающая сфинктер заднего прохода. Рефлекторная дуга от рецепторов прямой кишки замыкается в пояснично-крестцовом отделе спинного мозга. Она обеспечивает непроизвольный компонент акта дефекации. Произвольный компонент осуществляется при участии коры большого мозга, центров продолговатого мозга и гипоталамуса (рис. 8.17).
Из спинального центра дефекации парасимпатические нервные волокна в составе тазовых нервов тормозят тонус сфинктеров и усиливают моторику прямой кишки, стимулируя акт дефекации. Симпатические нервные волокна повышают тонус сфинктеров и тормозят моторику прямой кишки.
8.9.6.
 |
Микрофлора пищеварительного тракта
Пищеварительный тракт человека и животных «заселен» микроорганизмами. В одних отделах тракта в норме их мало или почти нет, в других — очень много. Макроорганизм и его микрофлора составляют единую динамичную систему. Динамичность эндоэкологического микробного биоценоза пищеварительного тракта определяется количеством поступающих в него микроорганизмов (за сутки перорально поступает около 1 млрд микробов), интенсивностью их размножения и гибели в пищеварительном тракте и выведения из него в составе кала (в норме выделяется за сутки 1012—1014 микроорганизмов).
Каждый отдел пищеварительного тракта имеет характерное для него количество и набор микроорганизмов. Их число в полости рта, несмотря на бактерицидные свойства слюны, велико (107—108 клеток в 1 мл ротовой жидкости). Содержимое желудка здорового человека натощак из-за бактерицидных свойств желудочного сока часто бывает стерильным; нередко обнаруживается и относительно большое число микроорганизмов (до 103 в 1 мл содержимого), проглатываемых со слюной. Примерно таково же число их в двенадцатиперстной и начальной части тощей кишки. В содержимом подвздошной кишки микроорганизмы обнаруживаются регулярно: в среднем 106 в 1 мл содержимого. В содержимом толстой кишки число бактерий максимальное.
Микрофлору кишечника делят на три группы:
• главная (бифидобактерии и бактероиды); составляет около 90 % всех микробов;
• сопутствующая (лактобактерии, эшерихии, энтерококки) — 10 % от общего их числа;
• остаточная (цитробактер, энтеробактер, протеи, дрожжи, клостридии, стафилококки, аэробные бациллы и др.) — менее I %.
Микроорганизмы, связанные со слизистой оболочкой кишечника, относятся к мукозной микрофлоре (Л/ -микрофлора), локализованные в полости кишки — к полостной (П-микрофлора). Соотношения между М- и П-микрофлорой динамичны. К внешним воздействиям М-микрофлора более устойчива, чем П-микрофлора. Количество П-микроорганизмов в направлении от желудка к толстой кишке нарастает более круто, чем М-форм; анаэробная микрофлора преобладает над аэробной.
За баугиниевой заслонкой резко изменяется и качество микрофлоры. Толстая кишка — это своеобразная микроэкологическая зона. В ней П-микрофлора представлена бактероидами, бифидобактериями, лактобактериями. В М-флоре толстой кишки доминируют бифидобактерии и лактобактерии. Общее число М-форм слизистой толстой кишки составляет 106, соотношение анаэробов к аэробам — 10:1. Максимальное число бактерий в фекалиях (10*°—1013 в 1 г), где они составляют до 30 % их массы.
Состав и количество микроорганизмов в пищеварительном тракте зависят от эндогенных и экзогенных факторов. К первым относятся влияния слизистой оболочки пищеварительного канала, его секретов и самих микроорганизмов, моторики. Ко вторым — характер питания, факторы внешней среды, прием антибактериальных препаратов. Экзогенные факторы влияют непосредственно и опосредованно через эндогенные факторы.
Существенны влияния на микрофлору функционального состояния пищеварительной системы. Перистальтика пищеварительного тракта обеспечивает транспорт микроорганизмов в составе химуса в каудальном направлении, что существенно в создании проксимодистального градиента заселенности кишечника микроорганизмами. Баугиниева заслонка предотвращает поступление микроорганизмов с содержимым из толстой кишки в тонкую.
Велика роль в формировании микрофлоры пищеварительного тракта пищеварительных секретов. Слюна имеет мурамидазу (лизоцим), которая определяет бактерицидные свойства этого секрета. Желудочный сок за счет соляной кислоты и других факторов обладает бактерицидностью, что существенно влияет на число и состав микрофлоры кишечника. Она зави-



|
Рис. 8.18. Потоки веществ из желудочно-кишечного тракта во внутреннюю среду организма в норме (А) и при недостаточности переваривания и всасывания нутриентов пищи (Б) (по А.М. Уголеву, с дополнениями).
сит и от поступления в тонкую кишку панкреатического, кишечного секретов и желчи. Эти влияния не только прямые, но и опосредованные. Так, снижение содержания нутриентов в химусе лишает микроорганизмы необходимых им питательных веществ. Гидролизаты белков и жиров ингибируют развитие ряда микроорганизмов. Таким действием обладают лейкопе- дез и свободные желчные кислоты, а также выделяемые в составе пищеварительных секретов иммуноглобулины, С-реактивный белок, лактоферрин.
Стабильность, сбалансированность и адекватность питания важны в стабилизации эубиоза человека.
Нормальная микрофлора — эубиоз — выполняет ряд важнейших для макроорганизма функций. Она участвует в формировании иммунобиологической реактивности организма, предохраняет макроорганизм от внедрения и размножения в нем патогенных микроорганизмов. Кишечная микрофлора синтезирует витамины К и группы В, другие вещества, важные для организма. В их числе физиологически активные амины (кадаверин, октамин, гистамин и др.), незаменимые аминокислоты, углеводы, жиры. Микрофлора обладает способностью морфокинетических воздействий на пищеварительный тракт, особенно на его слизистую оболочку. Ферменты бактерий расщепляют целлюлозу, гемицеллюлозу и пектины, непереваренные в тонкой кишке. Это симбионтный тип пищеварения, обеспечивающий поток из кишки вторичных нутриентов (рис. 8.18). Недостаточность собственного пищеварения увеличивает количество нутриентов, утилизируемых микрофлорой. Она количественно и качественно изменяется, что приводит ко многим патологическим последствиям.
Микроорганизмы утилизируют непереваренные питательные вещества, образуя при этом ряд веществ, которые всасываются из кишечника и включаются в обмен веществ организма. Микрофлора существенно влияет на печеночно-кишечную циркуляцию компонентов желчи и через них на 442 деятельность печени. С участием микрофлоры кишечника происходит обмен белков, фосфолипидов, желчных и жирных кислот, билирубина, холестерина.
Пищеварительные секреты, выполнив свою физиологическую роль, частично разрушаются и всасываются в тонкой кишке, а часть их поступает в толстую кишку, где они подвергаются действию микрофлоры. С участием микрофлоры инактивируются многие ферменты. Микроорганизмы принимают участие в разложении желчных кислот, ряда органических веществ с образованием органических кислот, их аммонийных солей, аминов и др.
8.10. ФУНКЦИИ ПЕЧЕНИ
Анатомическое положение печени на пути крови, несущей питательные и иные вещества от пищеварительного тракта, особенности строения, кровоснабжения, лимфообращения, специфика функций гепатоцитов определяют функции этого органа. Выше описана желчеотделительная функция печени, но она не единственная.
Важна барьерная функция, состоящая в обезвреживании токсичных соединений, поступивших с пищей либо образовавшихся в кишечнике за счет деятельности его микрофлоры, лекарств, всосавшихся в кровь. Химические вещества обезвреживаются путем их ферментативного окисления, восстановления, метилирования, ацетилирования, гидролиза (1-я фаза) и последующей конъюгации с рядом веществ — глюкуроновой, серной и уксусной кислотами, глицином, таурином и др. (2-я фаза). Не все вещества обезвреживаются в 2 фазы. В одну из них или без изменений выводятся в составе желчи и мочи растворимые конъюгаты. Инактивация токсичного аммиака происходит за счет образования мочевины и креатинина. Микроорганизмы обезвреживаются в основном путем фагоцитоза и лизиса.
Печень принимает участие в инактивации ряда гормонов (глюкокортикоиды, альдостерон, андрогены, эстрогены, инсулин, глюкагон, ряд гастроинтестинальных гормонов) и биогенных аминов (гистамин, серотонин, катехоламины).
Экскреторная функция печени выражается в выделении из крови в составе желчи большого числа веществ, обычно трансформированных в печени, что является ее участием в обеспечении гомеостаза.
Печень участвует в обмене белков', в ней синтезируются белки крови (весь фибриноген, 95 % альбуминов, 85 % глобулинов), происходит дезаминирование и переаминирование аминокислот, образование мочевины, глутамина, креатина, факторов свертывающей и противосвертывающей систем крови. Желчные кислоты влияют на транспортные свойства белков крови.
Печень участвует в обмене липидов: в их гидролизе и всасывании, синтезе триглицеридов, фосфолипидов, холестерина, желчных кислот, липопротеидов, ацетоновых тел, окислении триглицеридов. Велика роль печени в обмене углеводов: процессах гликогенеза, гликогенолиза, включении в обмен глюкозы, галактозы и фруктозы, образовании глюкуроновой кислоты.
Печень участвует в эритрокинетике, в том числе в разрушении эритроцитов, деградации гема с последующим образованием билирубина.
Важна роль печени в обмене витаминов, особенно жирорастворимых А, D, Е, К, всасывание которых в кишечнике идет с участием желчи. Ряд витаминов депонируется в печени и высвобождается по мере их метаболиче-
ской потребности (A, D, К, С, РР). Депонируются в печени микроэлементы (железо, медь, марганец, кобальт, молибден и др.) и электролиты.
Кишечно-печеночная циркуляция желчных кислот важна не только в гидролизе и всасывании липидов, но и в других процессах. Они являются регуляторами холереза и выделения в составе желчи холестерина, желчных пигментов, активности печеночных цитоферментов, влияют на транспортную активность энтероцитов, ресинтез в них триглицеридов, регулируют пролиферацию, передвижение, апоптоз и отторжение энтероцитов с кишечных ворсинок.
Регуляторное влияние желчи распространяется на секрецию желудка, поджелудочной железы и тонкой кишки, эвакуаторную деятельность гастродуоденального комплекса, моторику кишечника, реактивность органов пищеварения по отношению к нейротрансмиттерам, регуляторным пептидам и аминам.
Нормальное содержание в крови желчных кислот поддерживает и стимулирует физиологические и биохимические процессы. Они угнетаются при повышении концентрации желчных кислот в крови и затем проявляется их токсическое действие.
8.11. ПИЩЕВАРИТЕЛЬНЫЕ ФУНКЦИИ
И ДВИГАТЕЛЬНАЯ АКТИВНОСТЬ ЧЕЛОВЕКА
8.11.1. Влияние гипокинезии
Повседневная недостаточная физическая активность человека или ее минимизация в условиях специфической трудовой деятельности в ограниченном пространстве (космические и подводные корабли), при госпитальном постельном режиме, инвалидизация в связи с патологией опорно-двигательного аппарата, сердечно-сосудистой системы, центральных нарушений движений вызывает комплекс адаптационных перестроек в деятельности пищеварительной системы, названный К.В. Смирновым (1990) «гипокинетическим синдромом пищеварительной системы». По его мнению, данный синдром является результатом дефицита мышечной деятельности, приводящим к резкому уменьшению энергетических затрат организма, снижению активации ЦНС от рецепторов двигательного аппарата, уменьшению объемных параметров системной и регионарной гемодинамики с характерным снижением кровоснабжения органов пищеварения, застоем крови в них.
Важнейшим проявлением «гипокинетического синдрома пищеварительной системы» является гиперсекреция желудочных желез — увеличение секреции НС1 и пепсиногена в межпищеварительный период (натощак) и в ответ на стимуляцию секреции желудочных желез, что связывают с увеличением концентрации гастрина и Са2+ в крови. Секреция желудочной слизи при этом уменьшается. Такие изменения секреции — повышение агрессивных свойств желудочного сока и снижение защищенности слизистой от его действия — выступают как фактор риска язвенной болезни. Для описываемого синдрома характерны снижение моторной активности желудка, повышение тонуса пилорического сфинктера и замедление эвакуации из желуд








