 2014-02-02
2014-02-02 1322
1322Лекция 1. Развитие древостоев
О лесе известно как сложном образовании природы, явлении физико-географическом и биологическом. Лес изучается многими науками и достаточно давно, но осталось еще немало загадок и не открытых закономерностей.
Число сформулированных биоэкологами популяционных закономерностей очень высоко – до 20, а быть может, и значительно больше (Реймерс, 1994, с. 78). Причем многие из них взаимодополняют друг друга. Так, «закон популяционного максимума Ю. Одума» конкретизируют теория лимитов популяционной численности Х.Андреварты–Л.Бирча» и «теория биоценотической регуляции численности популяции К.Фридерихса» (Реймерс, 1994, с. 79). В сообществах древесных растений первые закономерности были выявлены еще в 19 веке, однако до сих пор пока нет общей теории онтогенеза древесных растений.
В общем виде влияние численности популяции на ее продуктивность было сформулировано А.А.Урановым (Уранов, 1960): «Наиболее доступный способ количественного выражения жизненности популяции вида состоит в определении количества органического вещества, производимого популяцией на единице территории за год. Это – мера мощности популяции. Она зависит от количества особей и возрастает по мере увеличения численности до некоторого предела – оптимальной численности и уменьшается при увеличении численности выше оптимальной».
В лесной таксации, лесных культурах и лесоводстве неоднократно предпринимались попытки установить численные параметры такой оптимальной численности, включая и теоретические расчеты (например, Стяжкин, 2005). Установление каких-либо закономерностей всегда наталкивается на мощное воздействие факторов среды – климата, колебаний инсоляции и осадков, богатства почвы и т.д., причем за весьма длительное время (до нескольких столетий). Выяснить законы развития и распада древостоев даже для одной древесной породы весьма непросто именно из-за длительности онтогенеза древесных видов.
В нашем представлении развитие простого одноярусного древостоя происходит следующим образом. Вначале появляется самосев или поросль, каждое из растений формирует крону и растения развиваются в молодые деревья. Постепенно кроны смыкаются и образуют полог древостоя. В пологе помещается некоторое ограниченное количество фотосинтезирующей поверхности: листьев, хвои, мелких веточек. Деревья растут и полог движется вверх, оставляя внизу отмирающие ветви и ослабленные растения. Деревья различаются по площади и объему кроны в десятки раз и эти различия определяют среди них лидеров и аутсайдеров. Лидеры определяются уже в самом раннем возрасте и деревья растут, просто увеличивая свои размеры и сохраняя свое ранговое положение в древостое (Маслаков,1980). Ранг дерева определяет именно величина кроны: чем она больше, тем прочнее положение дерева в древостое. В классическом лесоводстве деревья по степени развития кроны традиционно разделяют на 5 классов (классы Крафта).
На рост древостоев влияет множество факторов и причин. Одной из основных и решающих является конкуренция между растениями, влияние которой наиболее сильно начинает сказываться с началом смыкания крон (Петров, 1974; Кайрюкштис, Юодвалькис, 1976; Рогозин, 1980; Терехов, Усольцев, 2008). Онтогенез отдельных деревьев, а тем более древостоев достаточно сложен и весьма продуктивным для его описания оказался подход, предложенный Г.С.Разиным (Шинкаренко, Дзедзюля,1983).
Разработка данного подхода началась с выяснения вопроса изменчивости класса бонитета насаждений с возрастом. Для этого были использованы данные точных и продолжительных наблюдений в сосновых, еловых и березовых древостоях (всего 145 пробных площадей) в лесной опытной даче Тимирязевской сельхозакадемии за 100-летний период (Итоги работ…, 1964). По этим данным были построены линии роста древостоев и определены классы бонитета с точностью до десятых долей класса. Анализ линий роста у 85 сосновых древостоев показал, что за 20- летний период стабильный бонитет был обнаружен только у 23% древостоев, 72% меняли его на 1 класс, а у 5% древостоев он изменился на 2 класса; за 40 лет стабильный бонитет был у 35% древостоев, 40% меняли его на 1 класс, 20% - на 2 класса, а у 5% древостоев он изменился на 3-4 класса (!). У других пород такие же сильные изменения в росте происходили во всех возрастных периодах [7-10].
Впервые об изменении линий роста древостоев сообщалось еще в начале 20 века у Флюри и Гуттенберга, ссылаясь на которых Н.В.Третьяковым приводится пример, когда ель в 50 лет на трех участках имела высоту 13,5; 16,5 и 19,5 м, а в возрасте спелости таксировалась одним и тем же классом бонитета (Третьяков, 1937). Данные удивительные факты на протяжении многих десятилетий не анализировались должным образом. Даже в специальном обзоре работ, посвященном прогнозированию роста древостоев (Свалов, 1978) о причинах изменения хода роста древостоев упомянуто вскользь; по-видимому, причиной такого ухода от анализа неудобных фактов было стремление модифицировать и приспособить шкалу бонитетов М.М.Орлова для множества условий и пород, с целью ее использования для группировки древостоев при составлении таблиц хода роста древостоев. Тем не менее, Н.Н.Свалов все-таки делает вывод о том, что изменения классов бонитета с изменением возраста насаждений показали несовершенство бонитетной шкалы М.М.Орлова и ее пригодность для таксации древостоев только в статике (Свалов, 1978).
Этот вывод ключевой в понимании сущности вопроса о прогнозировании роста древостоев. Шкала М.М.Орлова «пригодна для таксации только в статике», то есть «здесь и сейчас» из чего логически вытекает вывод о ее непригодности для прогноза роста как в будущее, так и в прошлое. И если по ней подбирают возрастной ряд для составления ТХР, то по такой ТХР точно также будет невозможен прогноз роста на будущее. Парадоксально, но именно для такой цели (прогноза роста) и составляют таблицы хода роста древостоев!
Данные выводы весьма категоричны. В классическом лесоводстве и таксации леса оценка условий роста по классам бонитета прочно прижилась из-за простоты ее определения и каждый раз при обсуждении этого вопроса именно эта простота и унификация шкалы бонитетов перевешивает все другие аргументы против ее использования для составления ТХР.
Безусловно, для унифицированной оценки производительности условий местообитания нужны какие-то критерии. Вместо классов бонитета это может быть «верхняя высота», а также «элементарный запас» древостоев для каждой породы по отдельности, но это требует специального обсуждения. В конце концов, давно используются в таксации тип условий местопроизрастания (ТУМ), который не меняется во времени, адекватно отражает многообразие условий и может служить для группировки древостоев при составлении таблиц хода роста древостоев.
В середине 20 века составление ТХР основных лесообразующих пород было весьма актуальной задачей и об этом мы уже говорили выше в разделе 3. В настоящее время приняты две шкалы бонитетов: для семенных и порослевых насаждений; они унифицируют оценки роста для всех лесных пород в нашей стране и используются для оценки производительности условий местопроизрастания.
Здесь необходимо пояснить, что понятие бонитета насаждений было введено М.М.Орловым еще в 1911 г. в качестве косвенной оценки условий местообитания насаждений, их «добротности». Бонитет определяют по возрасту и средней высоте древостоя, с градацией высот между классами, например в 40 лет, через 2,5 м или с 20%-ным увеличением высоты от класса к классу. То есть в современном понимании оценка абиотических факторов питания подменяется оценкой биоты (фитоценоза) по успешности ее роста в высоту. Но если эта оценка меняется за 40 лет на 2-3 градации бонитета или до 60%, то следует ли ее применять для оценки производительности условий местопроизрастания, которые на самом деле не менялись?
Ответы на эти неудобные вопросы не получили иного объяснения кроме рабочей гипотезы Г.С.Разина о том, что изменения в ходе роста древостоев вызваны главным образом различиями в их начальной густоте. С этого предположения, собственно, и начались исследования в направлении, которые закончились открытием нижеследующих закономерностей.
В серии работ (Разин, 1965, 1977, 1979, 1980, 1988, 2009), с их обобщением в статьях (Разин, Рогозин, 2009, 2010а, б) а также в настоящей книге применительно к древостоям культур показано, что рост, развитие и возрастная динамика древостоев подчиняются некоторым всеобщим биологическим законам. В древесных насаждениях эти законы имеют специфические особенности, которые заключаются в следующем.
1. Одноярусные древостои всех пород, составов, различной начальной густоты и неравномерности расположения деревьев имеют своей целью общее стремление всех деревьев яруса к достижению предельных линейных, площадных и объемных показателей в пределах сообщества. В результате почти любой древостой однажды в жизни становится сомкнутым и достигает индивидуального предельного состояния по этим показателям.
2. Достижение предельных показателей происходит в зависимости от густоты растений в сообществе и может быть сформулировано в виде «Закона динамики суммы горизонтальных проекций крон деревьев в простых древостоях», который показан в виде нескольких линий зависимости на рисунке 4.1а и состоит в следующем:
- чем больше начальная густота, тем раньше и при меньшей средней высоте древостои становятся сомкнутыми с предельной суммой площадей горизонтальных проекций крон деревьев и тем раньше и быстрее они снижают ее (размыкаются);
- чем меньше начальная густота, тем позже и при больших средних высотах древостои достигают предел сомкнутости и предел суммы площадей горизонтальных проекций крон деревьев, тем медленнее они снижают их и дольше остаются сомкнутыми и устойчивыми.
Открытый для площадных показателей закон проявляет себя аналогичным образом в показателях объема. С некоторым запаздыванием по сравнению с сомкнутостью (на 10 – 25 лет) древостои точно также достигают предельного состояния по сумме объемов крон и имеют индивидуальные линии своего развития по этому показателю в зависимости от начальной густоты (рис 1). Впечатляют полученные различия в суммах объемов крон, достигающие 3-5 раз. Это приводят к различиям в объемах фотосинтезирующего аппарата, что обуславливает и разную производительность древостоев на таких линиях развития.
Интерпретация в биологическом плане обнаруженных законов выглядит следующим образом. Возникшие одноярусные сообщества древесных растений различной густоты и неравномерности расположения деревьев обладают сильнейшим изначальным природным свойством (способностью) у каждого члена сообщества, которое можно назвать целеполаганием или «целью» – стремлением к достижению индивидуальных пределов по протяженности (высоте, длине, толщине), а также по площади и по объему.
В древостое как сообществе изложенные «цели» появляются в следующем порядке: а) возникает сообщество индивидуумов, каждому из которых природой дана способность увеличивать размеры во времени и пространстве; б) сообщество, состоящее из устремленных в этих направлениях деревьев, становится обладателем суммы этих свойств в виде некоторой предельной цели.
Важно отметить, что при любых высотах и возрастах в ельниках бывают состояния с предельной сомкнутостью. При средней высоте 1 м сумма площадей крон деревьев может быть в 2,6 раза больше площади поверхности, на которой они расположены (предельная сомкнутость 2,60) и кроны деревьев в предельном случае заходят в кроны второго и даже третьего дерева по соседству. При высоте 20 м сомкнутость может иметь предельное значение 1,24. Достижение таких предельных значений воспринимается наблюдателем как древостой с почти полным отсутствием растительности в напочвенном покрове; тип леса определить затруднительно и условно он может быть назван «мертвопокровным».
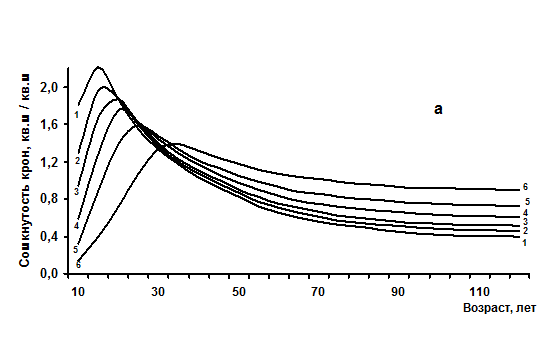
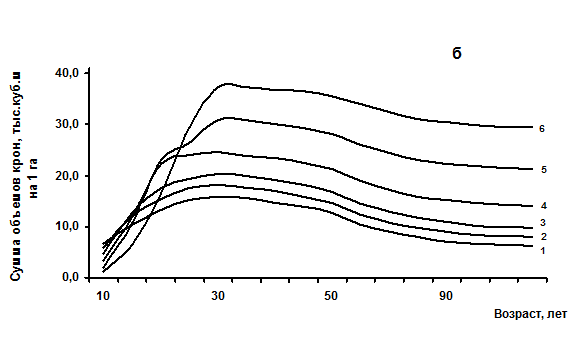
Рис. 1 – Возрастная динамика сомкнутости крон (а) и объемов крон (б) одноярусных еловых древостоев с различной начальной густотой (в возрасте 10 лет), тыс. шт/га: 1 - 67; 2 – 22; 3 – 11,5; 4 – 7; 5 – 4,5; 6 – 1,8 тыс. шт/га.
В интегрированном виде описанные выше законы могут быть объединены во всеобщий закон естественного роста, развития и жизни одноярусных древостоев, или сокращенно, закон развития одноярусных древостоев, который в кратком изложении выглядит следующим образом: «Чем больше начальная густота древостоев, тем меньшими в них по размерам оказываются со временем все деревья; густые древостои растут хуже, менее производительны, менее устойчивы к стрессам и быстрее распадаются из-за действия внутренних причин своего развития» (Разин, Рогозин, 2010, 2011).
Следствием данного закона является перегруппировка древостоев по мере их развития по всем таксационным показателям: сомкнутости, полноте, сумме объемов крон деревьев, запасам древесины, общей производительности и классам бонитета Эти изменения приводят к тому, что динамика абсолютной и относительной полноты также зависит от густоты: чем больше начальная густота, тем раньше достигает своих пределов полнота, тем быстрее она снижается и остается далее всегда меньшей, чем в более редких древостоях. Следствием являются и постоянные изменения рангов таксационных показателей древостоев по мере их развития. (Разин, Рогозин, 2010).
В спелых древостоях причиной низкой полноты чаще всего является прохождение ими состояния предельной сомкнутости ввиду большой начальной и последующей густоты их в молодости. Сами древостои недостаточно регулируют густоту и поэтому нуждаются в ее оптимизации. В наиболее перегущенных из них нужны разреживания с самого раннего возраста. При регулярном разреживании происходит переход древостоев на более производительные линии динамики. Это дает возможность революционного изменения способов выращивания древостоев на основе оптимальных программ регулирования текущей густоты (Разин, 1981, 1982, 1989).
В частности, при увеличении начальной густоты выращивания культур ели от 0,8 до 8,3 тыс. шт./га к возрасту 60-70 лет класс бонитета понижается на 1,2 единицы или на 18-20% по высоте (Разин, Рогозин, 2009, 2010). Множество других отличий и преимуществ редких культур перед густыми культурами были приведены в разделе 3.
В соответствии с обнаруженным законом можно однозначно утверждать, что начальная густота древостоев и возникающая конкуренция между растениями в решающей степени определяют всю последующую жизнь насаждения и чем больше густота, тем сильнее меняется рост и развитие как отдельных деревьев, так и древостоев в целом.








