 2014-02-03
2014-02-03 4065
4065Биоэнергетические функции митохондрий
Нарушение функций митохондрий при тканевой гипоксии
Нарушение биоэнергетических функций митохондрий - одно из наиболее ранних проявлений повреждения клеток.
Биоэнергетические функции митохондрий
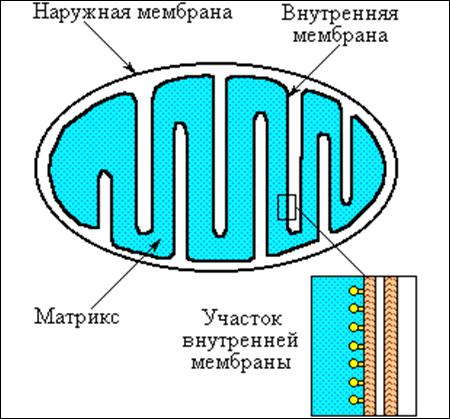 Митохондрии - это везикулярные структуры, образуемые наружной и внутренней мембранами (см. рис. 1). Внутренняя мембрана образует складки, или кристы, окружающие матрикс. На складках внутренней мембраны видны грибовидные выросты - это H+-АТФаза, или АТФ-синтаза, о которой будет сказано позже. Внутренняя мембрана митохондрий непроницаема для большинства ионов, включая ионы водорода, натрия, калия, хлора. Но она, как и другие биологические мембраны проницаема для воды; поэтому объем митохондрий зависит от концентрации ионов внутри и снаружи. В норме концентрации частиц по обе стороны митохондриальной мембраны равны, при патологии концентрация внутри обычно повышается и митохондрии "набухают", к чему мы ещё вернемся.
Митохондрии - это везикулярные структуры, образуемые наружной и внутренней мембранами (см. рис. 1). Внутренняя мембрана образует складки, или кристы, окружающие матрикс. На складках внутренней мембраны видны грибовидные выросты - это H+-АТФаза, или АТФ-синтаза, о которой будет сказано позже. Внутренняя мембрана митохондрий непроницаема для большинства ионов, включая ионы водорода, натрия, калия, хлора. Но она, как и другие биологические мембраны проницаема для воды; поэтому объем митохондрий зависит от концентрации ионов внутри и снаружи. В норме концентрации частиц по обе стороны митохондриальной мембраны равны, при патологии концентрация внутри обычно повышается и митохондрии "набухают", к чему мы ещё вернемся.
Рис. 1. Схема строения митохондрий
Окислительное фосфорилирование
Митохондрии осуществляют важнейшую для клеточной биоэнергетики реакцию фосфорилирования АДФ с образованием АТФ за счет энергии окисления органических соединений, служащих субстратами окисления, молекулярным кислородом. Конечная стадия этого процесса - перенос электронов от восстановленных пиридиннуклеотидов и сукцината на молекулярный кислород осуществляется по системе переносчиков электрона, которая в совокупности называется дыхательной цепью (см. рис. 2).
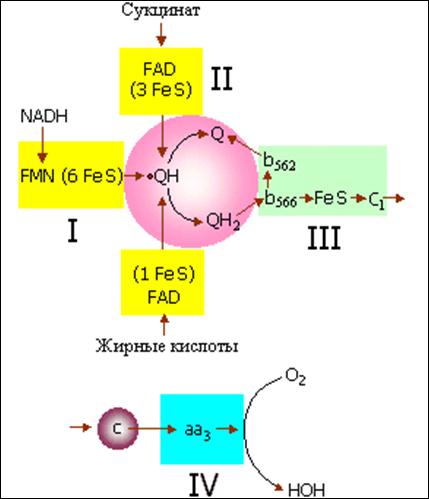
Рис.2 Дыхательная цепь митохондрий (схема). Римскими цифрами обозначены дыхательные комплексы, на которые мембраны митохондрий впервые разделил Дэвид Грин. Строчными буквами обозначены цитохромы, остальные сокращения общеприняты в биохимии.
При переносе электронов по дыхательной цепи происходит высвобождение энергии, величина которой (в электрон-вольтах) равна разности стандартных восстановительных потенциалов двух реагирующих ред-окс пар. Эти стандартные потенциалы приведены на рис. 3.
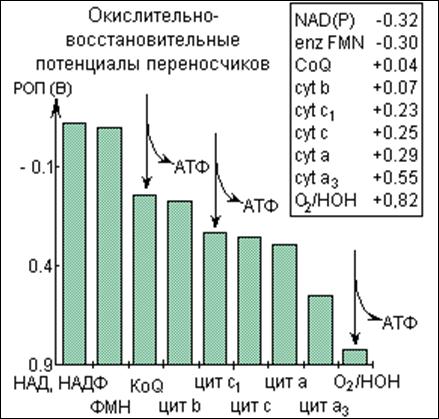 Рис.3. Восстановительные потенциалы переносчиков электронов в дыхательной цепи митохондрий
Рис.3. Восстановительные потенциалы переносчиков электронов в дыхательной цепи митохондрий
На участках с большим перепадом энергии осуществляется синтез АТФ (показано стрелками). Эти участки называются точками сопряжения.
Английский биохимик Петер Митчелл сначала предположил, а потом показал, что одновременно с переносом электронов по дыхательной цепи митохондрий в определенных ее звеньях, называемых точками сопряжения, происходит перенос протонов из матрикса в окружающую среду. При этом энергия окислительно-восстановительной реакции DE превращается в энергию перенесенных протонов.
Энергия одного моля иона в данной среде называется, как известно, электрохимическим потенциалом. Разность электрохимических потенциалов протона между двумя водными фазами внутри и вне митохондрий описывается уравнением:
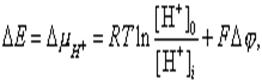 (1)
(1)
где R - газовая постоянная, T - абсолютная температура, [H+]o и [H+]i - концентрации ионов водорода вне и внутри матрикса, соответственно, F - число Фарадея, Dj - разность потенциалов между окружающей средой и матриксом.
Петер Митчелл в качестве единицы энергии использовал электрон-вольты, в результате чего уравнение (1) несколько трансформируется:

Суммарная энергия окислительно-восстановительной реакции, превращенная в разность электрохимических потенциалов ионов водорода, была названа П. Митчеллом протон-движущей силой (PMF - proton motive force), по аналогии с электродвижущей силой в гальванической батарее. Заменив натуральный логарифм десятичным, легко найти величину протон-движущей силы, зная разность pH (DpH) и разность потенциалов (Dj) между средой и матриксом при комнатной температуре; выраженная в милливольтах она будет равна:
PMF (мВ) = 60 (мВ) · DpH + Dj (2)
В митохондриях основной вклад в эту сумму вносит мембранный потенциал, который в присутствии субстрата и кислорода составляет около 170-180 мВ. Созданная работой дыхательной цепи разность потенциалов DmH+ может быть использована для синтеза АТФ или переноса ионов в митохондрии. Синтез АТФ осуществляется благодаря работе АТФ-синтазы, которая представляет собой протонную АТФазу (H+-АТФазу), работающую как бы "задним ходом" (см. схему на рис. 4).
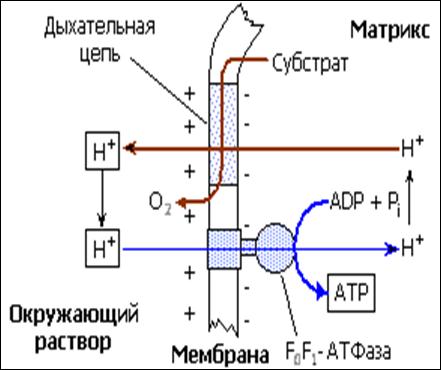
Рис.4. Схема окислительного фосфорилирования, согласно хемоосмотической теории Митчелла. При переносе электронов по дыхательной цепи в каждой точке сопряжения происходит перенос протонов через мембрану (один электрон – один протон) из матрикса наружу. Создаваемая разность элетрохимических потенциалов протона (H+) служит движущей силой для работы АТФ-синтазы (F0F1-АТФаза). Электродиффузионный перенос протонов из окружаюшей среды в матрикс "проворачивает колесо фермента" и происходит синтез АТФ. Рисунок взят из учебника Ленинждера (т.2, стр. 531, 1985).

