 2014-02-10
2014-02-10 4096
4096Митохондрии – органеллы клетки обычно вытянутой мешочковидной формы размерами в длину 4 – 7 мкм и в диаметре 0,5 - 2,0 мкм с эластичной двойной мембраной и студнеобразным матриксом. Внутренняя мембрана образует ряд выростов (крист), разделяющих полость митохондрии (мaтpuкc) на ряд отсеков, соединяющихся между собой. Между мембранами в так называемом перимитохондриальном пространстве находится жидкость (энхилема). Матрикс митохондрии содержит большое количество ферментных систем, принимающих участие в процессе дыхания. На кристах также расположена часть дыхательных ферментов. Мембраны митохондрии содержат большое количество белка (65 %) и липидов (35 %). Липиды мембран митохондрий очень богаты ненасыщенными жирными кислотами.
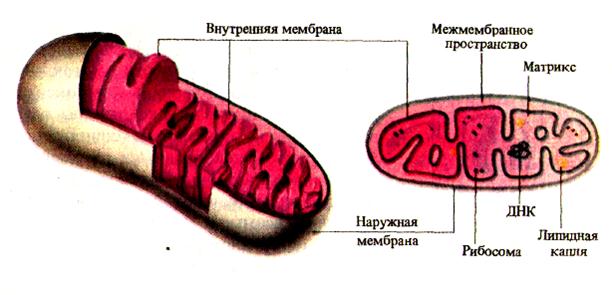
Строение митохондрии
В живой клетке постоянно происходит обновление митохондрий, время индивидуальной жизни которых колеблется от нескольких часов до нескольких суток. Митохондрии образуются не только из отдельных инициалей, но и путем деления сформировавшихся митохондрий перетяжками или почкованием.
Митохондрии являются энергетическими центрами клетки. В них происходит окисление органических веществ в процессе дыхания до СО2, и Н2О и запасание энергии в макроэргических связях молекулы АТФ.
Пероксисомы и глиоксисомы. В растениях присутствуют округлые органоиды диаметром 0,2 - 1,5 мкм, ограниченные элементарной мембраной и содержащие гранулярный матрикс умеренной электронной плотности. Они получили название микротел. В некоторых микротелах обнаруживается белковый кристаллоид, состоящий из трубочек диаметром около 6 нм. Количество микротел в клетке близко к числу митохондрий. В клетках растений обнаружены два типа микротел, выполняющих различные физиологические функции: пероксисомы и глиоксисомы.
Пероксисомы многочисленны в клетках листьев, где они тесно связаны с хлоропластами. В них окисляется синтезируемая в хлоропластах в ходе фотосинтеза гликолевая кислота и образуется аминокислота глицин, которая в митохондриях превращается в серии. В листьях высших растений пероксисомы участвуют в фотодыхании,
Глиоксисомы появляются при прорастании семян, в которых запасаются жиры, и содержат ферменты, необходимые для превращения жирных кислот в сахара: системы (b-окисления жирных кислот и глиоксилатный цикл). При работе ферментных систем пероксисом и глиоксисом образуется пероксид водорода, который разрушается содержащейся в этих органоидах каталазой.
Сферосомы. Это сферические, сильно преломляющие свет образования диаметром 0,5 мкм. Они содержат липиды и поэтому их называют также липидными каплями (олеосомами). В сферосомах обнаружены такие ферменты, как липаза и эстераза. В них хранятся запасы липидов клетки. При прорастании семян, запасающих жиры, сферосомы функционируют в комплексе с глиоксисомами в процессах глюконеогенеза.
Вакуолярная система. Вакуоль - типичный органоид растительной клетки. В меристематических клетках вакуоли представлены мелкими пузырьками; для зрелых клеток характерна большая центральная вакуоль. Вакуолярная система растений формируется несколькими путями. Из расширенных цистерн ЭР образуются провакуоли, слияние которых приводит к возникновению более крупных вакуолей и созданию вакуолярной мембраны - тонопласта, который таким образом является производным ЭР. Тонопласт может образовывать инвагинации, что приводит к включению в вакуоль участков цитоплазмы. Гидролитические ферменты, содержащиеся в возникшей вакуоли, расщепляют полимеры до низкомолекулярных веществ.
Важную роль в становлении вакуолей играют явления автофагии. Процесс начинается с окружения мембраной ЭР участка цитоплазмы (автофагическая вакуоль). Деятельность кислых гидролаз в замкнутом мембраной пространстве приводит к деградации полимерного содержимого и поступлению воды. Вакуоли, возникшие в результате автофагии, могут сливаться с вакуолями, образовавшимися иными путями.
Вакуолярный сок имеет сложный состав и включает органические вещества и минеральные соли. Помимо органических кислот, углеводов, аминокислот и белков, которые могут быть - вторично использованы в обмене веществ, клеточный сок содержит фенолы, таннины, алкалоиды, антоцианы, которые выводятся из обмена веществ клетки в вакуоль и таким путем изолируются от цитоплазмы. Большинство ферментов вакуолей - гидролазы с оптимумом активности при кислом рН, что позволяет рассматривать вакуоли растительных клеток в качестве вторичных лизосом. Кислотность вакуолярного сока 5,0 - 6,5 единиц рН, но может быть равной 1,0 (бегония) или 2,0 (лимон).
Вещества доставляются в вакуоль с помощью разнообразных систем транспорта, функционирующих в тонопласте. К ним относится АТР-зависимая Н + -помпа, выносящая ионы Н + из цитоплазмы в вакуоль (см. рис. 1.3). Ее деятельность обеспечивает поступление в вакуоль анионов органических кислот, Сахаров, а также вход и выход ионов К+. Переносчики, локализованные в тонопласте, обусловливают накопление в вакуолях аминокислот и других соединений.
Важно отметить, что вакуоль может служить местом отложения запасных белков (алейроновые зерна). Процесс вакуолизации - необходимое условие роста клеток растяжением.
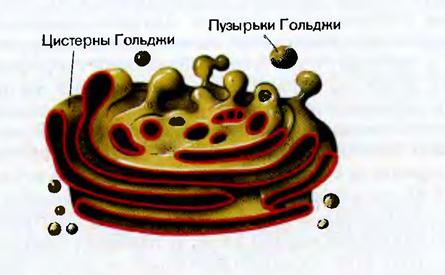
Аппарат Гольджи. В растительных клетках аппарат Гольджи (АГ) представлен диктиосомами, везикулами и межцистерными образованиями. Уплощенные цистерны - диктиосомы расположены пачками по несколько штук. Они ограничены мембраной толщиной 7 - 8 нм. На регенерационном полюсе АГ происходит новообразование диктиосом из мембран гладкого ЭР. На секреторном полюсе формируются секреторные пузырьки (везикулы), содержащие предназначенные для секреции вещества. В клетке растений содержатся от нескольких до сотен АГ.
В диктиосомах АГ образуются гликопротеины и гликолипиды и осуществляется накопление и мембранная «упаковка» соединений, необходимых для синтеза полимеров клеточной стенки и различных растительных слизей. С помощью везикул Гольджи углеводные компоненты доставляются к плазмалемме. Мембрана пузырьков встраивается в плазмалемму, способствуя ее росту и обновлению. Секретируемые вещества оказываются в клеточной стенке. Мембраны АГ являются связующим звеном между мембранами ЭР и плазмалеммой.
Эндоплазматический ретикулум. Эндоплазматический ретикулум (ЭР), или эндоплазматическая сеть (ЭС), представляет собой систему каналов, пузырьков и цистерн, ограниченную мембраной толщиной 5-6 нм. ЭР может содержать на своей поверхности рибосомы (гранулярный, или шероховатый, ЭР) или не содержать их (агранулярный, или гладкий, ЭР). Поверхность мембран ЭР в клетке больше других мембранных образований, а объем полостей может достигать 16% объема клетки. Эндоплазматическая сеть - очень лабильная структура. Неблагоприятные внешние воздействия (недостаток кислорода и др.) вызывают концентрические закручивания мембран ретикулума.
В мембранах ЭР локализованы редокс-цепи двух типов, с участием которых происходят детоксикация вредных для клетки соединений и превращение насыщенных жирных кислот в ненасыщенные. В гладком ЭР образуются углеводы, липиды, терпеноиды. В гранулярном ретикулуме синтезируются мембранные белки, ферменты, необходимые для синтеза полисахаридов клеточных стенок, структурный белок и ферменты клеточных стенок, другие секретируемые белки. По системе ЭР переносятся вещества внутри клетки. Он участвует также в межклеточных взаимодействиях у растений через плазмодесмы.
Рибосомы. Рибосомы осуществляют синтез белков - трансляцию матричной, или информационной, РНК (мРНК). На электронных фотографиях они выглядят округлыми частицами диаметром 20 - 30 нм. Каждая рибосома состоит из двух нуклеопротеиновых субъединиц. В цитоплазме растительных клеток находятся 80 S рибосомы, состоящие из 40 и 60 S субъединиц, в хлоропластах - 70 S рибосомы, а в митохондриях - 78 - 80 S рибосомы, отличные от цитоплазматических и хлоропластных. Субъединицы рибосом, образованные в ядрышке, поступают в цитоплазму, где происходит сборка рибосом на молекуле мРНК.
Рибосомы в цитоплазме могут быть свободными, прикрепленными к мембранам эндоплазматического ретикулума, к наружной мембране ядерной оболочки либо образуют полирибосомные (полисомные) комплексы. Полирибосомы возникают потому, что одну молекулу мРНК могут одновременно транслировать несколько рибосом. Полирибосомы разрушаются при воздействии на растения неблагоприятных факторов внешней среды (например, засухи, недостатка кислорода). В процессе синтеза белка, осуществляемого рибосомами, участвуют компоненты, образованные в ядре, ядрышке, в цитоплазме, митохондриях и хлоропластах.
Микротрубочки и микрофиламенты (цитоскелет). В наружном кортикальном слое цитоплазмы неделящихся растительных клеток локализованы микротрубочки. Их наружный диаметр 30 нм, внутренний - около 14 нм. Они ориентированы параллельно друг другу и перпендикулярно продольной оси клетки. При делении клетки микротрубочки составляют основу структуры веретена, пучки трубочек прикрепляются также к кинетохорам хромосом. У монадных водорослей микротрубочки входят в состав жгутиков, обеспечивая их подвижность. Все микротрубочки имеют единый план строения и состоят из глобулярного кислого белка тубулшш, субъединицей которого является димер, состоящий из двух глобулярных мономеров (a- и b-тубулинов). В микротрубочке димеры белка располагаются по спирали. Трубка образована 13 субъединицами тубулина. Цитоплазматические микротрубочки легко диссоциируют на субъединицы (разбираются) и собираются вновь. Для сборки микротрубочек благоприятен кислый рН среды, присутствие магния, GTP и АТР. Разборка ускоряется повышением концентрации Са2+ и низкой температурой.
В цитоплазме растительных клеток обнаружены также филаментные структуры, состоящие из немышечного актина. Это сократительный белок, сходный по молекулярной массе с актином мышц и близкий ему по аминокислотному составу. Он может находиться в мономерной (глобулярный, Г-актин) или в полимерной форме двойной спирали (фибриллярный, Ф-актин). Микрофиламенты актина взаимодействуют с микротрубочками кортикального слоя и плазмалеммой. Они участвуют в пространственной организации метаболических процессов, протекающих в растворимой фазе цитоплазмы, и служат основой ее двигательной активности.
Ядро. В растительной клетке ядро имеет диаметр порядка 10 мкм. Оно может быть сферическим, удлиненным или лопастным. Внутреннее содержимое ядра (нуклеоплазма) ограничено ядерной оболочкой, состоящей из двух элементарных мембран - внутренней и наружной. Ядерная оболочка пронизана порами диаметром 10-20 нм, через которые транспортируются нуклеиновые кислоты и белки. С наружной мембраной ядерной оболочки могут быть непосредственно связаны элементы эндоплазматического ретикулума. В нуклеоплазме локализованы участки хроматина, состоящего из комплекса ДНК, РНК и белков. В делящихся клетках хроматин организуется в хромосомы, число которых специфично для каждого вида растений. Количество ДНК в одном ядре постоянно для каждого вида (кроме случаев мейоза и полиплоидии). Геном растений отличается от генома животных большим количеством ДНК. В нуклеоплазме ядра содержатся также ферменты и кофакторы, необходимые для обеспечения процессов репликации и транскрипции ДНК, молекулы различных РНК, ферменты, фосфорилирующие и ацетилирую-щие ядерные белки, ферменты гликолиза и др.
Ядрышко отчетливо видно в ядре под световым и электронным микроскопами. Оно формируется на определенных участках ДНК, называемых ядрышковым организатором. В хроматине ядрышка находятся участки ДНК, ответственные за синтез рибосомальных РНК (рРНК).
В целом ядро является местом хранения генетической информации клетки и репликации ДНК. В нем происходит процесс транскрипции ДНК в РНК различных типов. В тесном взаимодействии с цитоплазмой ядро участвует в обеспечении экспрессии генетической информации и контролирует процессы жизнедеятельности клетки.

