 2014-02-12
2014-02-12 2972
2972КАРАГАНДИНСКАЯ ГОСУДАРСТВЕННАЯ МЕДИЦИНСКАЯ АКАДЕМИЯ
КАФЕДРА – МЕДИЦИНСКОЙ БИОФИЗИКИ И ИНФОРМАТИКИ
Лекция Формальное описание ионных токов в модели Ходжкина – Хаксли.
ПО ООD 012 МВ 1112 - МЕДИЦИНСКОЙ БИОФИЗИКЕ
ДЛЯ СПЕЦИАЛЬНОСТЕЙ: 051301 – «ОБЩАЯ МЕДИЦИНА»
051302 – «СТОМАТОЛОГИЯ
КУРС – 1
Продолжительность 50 минут
Составитель:
доцент, к.б.н.
____________ И.М. Риклефс
Караганда 2007 г.
Утверждена на заседании кафедры
Протокол №_____
от "____"__________200___г
Зав.кафедрой доцент ______________ Б.К. Койчубеков
Учебная цель лекции: Изучить физические и физико-химические механизмы генеза мембранного потенциала. Показать молекулярную основу происхождения мембранных потенциалов и их роль в проведении возбуждения. Обосновать роль изменения скорости передачи информации в изменении функционального состояния организма.
ПЛАН
1.ПОТЕНЦИАЛ ДЕЙСТВИЯ НЕРВНОГО ВОЛОКНА. ФОРМАЛЬНОЕ ОПИСАНИЕ ИОННЫХ ТОКОВ В МОДЕЛИ ХОДЖКИНА - ХАКСЛИ
2. ИОННЫЕ КАНАЛЫ КЛЕТОЧНЫХ МЕМБРАН
2.1. Независимость работы отдельных каналов.
2.2. Дискретный характер проводимости ионных каналов.
2.3.Зависимость параметров канала от мембранного потенциала.
2.4.Структура ионного канала.
1. ФОРМАЛЬНОЕ ОПИСАНИЕ ИОННЫХ ТОКОВ
В МОДЕЛИ ХОДЖКИНА - ХАКСЛИ
Данные, полученные методом фиксации потенциала, позволили произвести количественное описание ионных токов через мембрану и тем самым сформулировать математическую теорию нервного импульса. Основой создание математической модели ионных токов в мембране аксона кальмара стали опыты по разделению ионных токов возбужденного аксона.
Было показано, что если использовать два типа блокаторав – натриевого канала терродотоксин (ТТХ) и калиевого канала – тетраэтиламонний (ТЭА), то при регистрации в режиме фиксации потенциала на мембране то можно получить три типа кривых (слайд 1)
На основе опытов по фиксации напряжения на аксоне кальмара А.Ходжкин и А.Хаксли сформулировали ряд принципов:
1. Перенос Na+ и К+ осуществляется различными независимыми структурами;
2. Изменение токов являются следствием изменения проницаемости мембраны для ионов Na+ и К+;
3. Пропускная способность мембраны управляется электрическим полем.
Модель, предложенная Хаксли и Ходжкиным, предполагает, что проводимость мембраны для ионов Na+ и К+ регулируется некоторыми управляющими частицами, перемещающимися в мембране при изменениях электрического поля. Рассматривая потенциалозависимые частицы, регулирующие состояние калиевого канала Ходжкин и Хаксли выяснили, что если бы канал открывался в результате перехода слева направо только одной активирующей частицы, то проводимость ионных каналов менялась во времени по экспоненте, т.е. можно было бы записать:
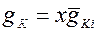 , (1)
, (1)
где 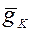 - максимальная проводимость для ионов типа K. Однако, наилучшее соответствие кинетической кривой калиевой проводимости в аксоне кальмара достигается при допущении, что для открывания калиевых каналов необходим приход к определенному участку мембраны четырех активирующих частиц.
- максимальная проводимость для ионов типа K. Однако, наилучшее соответствие кинетической кривой калиевой проводимости в аксоне кальмара достигается при допущении, что для открывания калиевых каналов необходим приход к определенному участку мембраны четырех активирующих частиц.
Пусть n обозначает вероятность того, что одна частица, управляющая К+-проницаемостью находится одновременно четыре частицы, равна n4.
Поэтому К+-проводимость мембраны в любой момент времени равна:
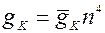 (2),
(2),
где - максимальная проводимость при сильной и длительной деполяризации мембраны, n4 – доля gK, которая включена в данный момент времени. Четвертая степень определялась эмпирически.
Частицы, управляющие проницаемостью К+-каналов, называются n-частицами.
Величина К+-проводимости, пропорциональная четвертой степени n, растет уже не по экспоненте, а с большим периодом задержки. Быстрое изменение величины n4 начинается лишь после того, как значение n превысит 0,7.
Значительно большие трудности возникли у Ходжкина и Хаксли при количественном описании переходящего повышения натриевой проводимости. Выбор метода описания зависел от принятия той или иной гипотезы о взаимоотношениях между процессами активации и инактивации натриевых каналов. Альтернатива, стоящая перед авторами, заключалась в следующем. Можно было принять, что инактивация представляет собой результат каких-то вторичных, возможно, химических изменений "активирующих" частиц, пришедших в район натриевого канала при деполяризации мембраны. В этом случае для формального описания кинетики gNa достаточно использовать одну переменную, подчиняющуюся дифференциальному уравнению второго порядка.
Другое объяснение падения gNa при продолжающейся деполяризации мембраны основано на предположении, что инактивация является совершенно независимым от активации процессом и что она обусловлена приходом в район натриевого канала особых "инактивирующих" частиц. При таком допущении изменения натриевой проводимости должны описываться двумя независимыми переменными, каждая из которых удовлетворяет уравнению первого порядка. Эта вторая альтернатива допускала более простое описание экспериментальных данных, и поэтому Ходжкин и Хаксли остановились на ней.
Кинетические кривые Na+-проводимости имеют более сложную форму (слайд 2).: проводимость нарастает до max-активация, а затем снижается – инактивация.
Изменение Na-проводимости, согласно второй альтернативе, предполагает наличие активирующих частиц m и инактивирующих h. Предполагают, что для открывания канала необходимо поступление в определенный участок мембраны трех m-частиц. Переход через мембрану одной инактивирующей частицы вызывает блокировку канала. Таким образом, изменения Na-проводимости описывают уравнением:
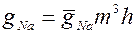 (3)
(3)
где 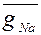 - максимальная Na-проводимость,
- максимальная Na-проводимость,
m – вероятность нахождения в заданном участке одной m-частицы,
h – вероятность того, что инактивирующая частица в этом участке отсутствует.
При наложении поля, перемещающего заряженные частицы с левой стороны мембраны на правую, m возрастает в пределах от 0 до 1, а h снижается в пределе до 0. Ходжкин и Хаксли создают систему уравнений описывающую, возникновение мембранных потенциалов(слайд 3).
После того как предложенная система уравнений была сформулирована, естественно возник вопрос, в какой мере она может быть применена для описания нормальной электрической активности волокна. "Было ясно, - писал Хаксли, - что использованные формулировки являются отнюдь не единственными возможными для описания опытов с фиксацией потенциала, и поэтому ни в коем случае нельзя было быть заранее уверенным, что те же уравнения смогут описать поведение мембраны и при нормальных условиях ее работы, когда ионные токи вызывают изменения мембранного потенциала вместо того, чтобы быть подавленными усилителем обратной связи. Мы поэтому рассчитали ответы нашей математической модели нервной мембраны на эквивалентный стимул вручную, а затем на ЭВМ".
На основе рассмотренной математической модели А. Ходжкину и А. Хаксли удалось с высокой точностью воспроизвести нервный импульс, генерируемый мембраной аксона в различных экспериментальных условиях. После незначительной модификации модели был имитирован процесс электрического возбуждения в миелинизированных нервных волокнах и соме нервных клеток.
Разделение трансмембранных токов на отдельные компоненты сыграло чрезвычайно важную роль для понимания механизмов электровозбудимости мембраны: охарактеризованы селективность, потенциалозависимость, кинетика включения и выключения внутримембранных путей – ионных каналов. В результате созданы общие представления о структурно-функциональной организации Na+ и К+- каналов, управляемых изменениями электрического поля: выделены функциональные элементы ионных каналов, ответственные за механизмы селекции, селективный фильтр, активации – "активные ворота" и инактивации –"инактивационные ворота".
Выяснилось, что к числу образований с "аномальным", с точки зрения классического варианта гипотезы Ходжкина механизмом генерации потенциалов действия наряду с упомянутыми клетками относятся волокна гладких мышц, миокардиальные клетки, многие нейроны моллюсков и позвоночных животных.

