2014-02-12
2014-02-12 2764
27645.3.1 Натрий – калиевый насос
Согласно современным представлениям, в биологических мембранах имеются ионные насосы, работающие за счет свободной энергии гидролиза АТФ. Это специальные системы интегральных белков – транспортные АТФазы. В настоящее время известны три типа электрогенных ионных насосов, которые осуществляют активный транспорт ионов через мембрану (рис.5.10).
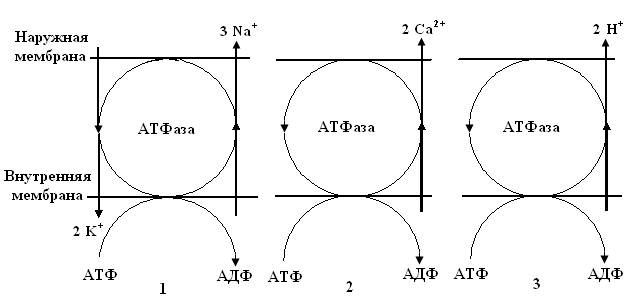
Рис.5.10.Виды ионных насосов: 1 – К+-Na+-АТФаза в цитоплазматических мембранах (К+-Na+- насос); 2 – Са2+-АТФаза (Са2+- насос); 3 – Н+-АТФаза в энергосопрягающих мембранах митохондрий и хлоропластов (Н+-насос).
Перенос ионов транспортными АТФазами происходит вследствие реакции сопряжения данного процесса с химическими реакциями распада АТФ, то есть за счет энергии метаболизма клеток. При работе К+-Na+-АТФазы за счет энергии, освобождающейся при гидролизе АТФ, в клетку переносится два иона калия и одновременно из клетки выкачиваются три иона натрия. Таким образом, создается повышенная по сравнению с межклеточной средой концентрация в клетке ионов калия и пониженная натрия.
Механизм работы ионных насосов подробно изучен на примере К+-Na+-АТФазы. (Рис. 5.11). Процесс работы данного фермента состоит в следующем (Е – фермент; Е1 и Е2 – соответствуют расположению активного центра фермента на внутренне и внешней поверхности мембраны соответственно; звездочкой обозначен активный комплекс).
1.Взаимодействие фермента с АТФ и образование активного комплекса на внутренней поверхности мембраны (эта реакция активируется катионами магния):
Е + АТФ → Е*АТФ.
2.Связывание комплексом трех ионов натрия:
Е*АТФ + 3 Na+ → [Е*АТФ]*Na3.
3.Фосфорилирование фермента с образованием АДФ:
[Е*АТФ]*Na3 → [E1 ~ P]* Na3 + АДФ.
4.Конформационное изменение фермента внутри мембраны:
[E1 ~ P]* Na3 → [E2 ~ P]* Na3.
5.Реакция ионного обмена натрия на калий, происходящая на внешней поверхности мембраны:
[E2 ~ P]* Na3 + 2 К+ → [E2 ~ P]* К2 + 3 Na+.
6.Обратное конформационное изменение комплекса с переносом ионов калия внутрь клетки:
[E2 ~ P]* К2 → [E1 ~ P]* К2.
7.Возвращение фермента в исходное состояние с освобождением ионов калия и неорганического фосфата (Р):
[E1 ~ P]* К2 → Е + Р + 2 К+.
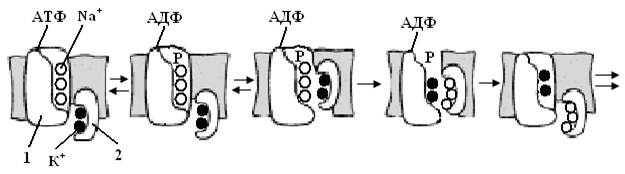
Рис.5.11.Рабочий цикл фермента К+-Na+-АТФазы (1 – большая субъединица фермента; 2 – малая субъединица; Р – фосфат.).
К+-Na+-АТФаза состоит из двух полипептидных цепей с молекулярной массой 84 000 и 5 700, которые формируют большую и малую субъединицы фермента. Белок прочно связан с фосфолипидами, полное удаление которых из мембран приводит к исчезновению АТФазной активности.
Катионы Na+ и К+ одновременно с разными субъединицами, между которыми возможен обмен катионами. Na+- связывающие центры находятся в большой субъединице фермента. Ионообменная полость открывается с внутренней поверхности мембраны, а внешний вход в эту полость (канал) закрыт гидрофобным контактом липидов и белков.
Гидролитический центр расположен на большой субъединице с внутренней стороны мембраны. В исходном состоянии ионообменные полости субъединиц могут заполняться катионами только из примембранных слоев. Эти полости из-за стерических затруднений могут вмещать три иона натрия и только два иона калия.
Конформационное состояние фермента определяет специфичность связывания ионов большой субъединицей. Для образования стабильного фосфорилированного комплекса необходимо, чтобы с большой субъединицей были связаны три иона натрия. Присоединение АТФ индуцирует конформационные изменения, в результате которых сродство большой субъединицы к Na+ значительно возрастает. Одновременно увеличивается отрицательный заряд ионообменной полости за счет близко расположенной фосфатной группы АТФ.
В результате ионного обмена с внутриклеточной средой оказываются связанными три иона натрия, а затем происходит фосфорилирование большой субъединицы. В результате последующих конформационных перестроек три иона натрия оказываются замкнутыми в ионообменной полости большой субъединицы. На этой стадии сродство большой субъединицы к натрию понижается, а к калию возрастает. Фермент переходит в стабильное состояние.
Дальнейшее протекание реакции связано с участием малой субъединицы. Субъединицы фермента могут незначительно смещаться друг относительно друга в результате теплового движения. Когда полость малой субъединицы совмещается с полостью большой, между ними происходит обмен катионами.
Замена трех катионов натрия на два калия в большой субъединице вновь приводит к возникновению конформационно неравновесного состояния. Уменьшение числа положительных зарядов в полости большой субъединицы сопровождается исчезновением большого активационного барьера. Этот барьер препятствует дефосфорилированию фермента и и отщеплению отрицательно заряженной фосфатной группы. При этом происходит открывание полости большой субъединицы во внутриклеточную среду. После диссоциации АДФ и Р фермент возвращается в исходное состояние.
Активный перенос ионов калия и натрия имеет большое значение при проведении нервного импульса. На транспорт этих ионов затрачивается энергия ∆G = 41,2 кДж/моль; примерно такая же энергия выделяется при гидролизе 1 моль АТФ до АДФ и фосфата: ∆G = − 36,0 кДж/моль.