 2014-02-12
2014-02-12 2021
2021Документационное обеспечение системы защиты информации
Механизм системы защиты информации не может успешно функционировать без соответствующего документационного обеспечения. Обычно предлагается пакет документов по системе защиты информации, который можно разделить на 3 группы:
А) документы закрепляющие право на коммерческую информацию;
Б) документы системы защиты информации;
В) документы по организации и ведению делопроизводства документов содержащих коммерческую тайну.
Ферменты (энзимы) – специфические белки, входящие в состав всех клеток и тканей живых организмов, играющие роль биологических катализаторов.
Доказательства белковой природы ферментов.
1. Инативация ферментов при нагревании. Инактивация ферментов совпадает с денатурацией белка. Ферменты разрушаются также под действием минеральных кислот, щелочей, солей, алкалоидов, при облучении рентгеновскими и ультрафиолетовыми лучами.
2. Электрохимические свойства ферментов.
a. Изоэлектрическая точка ферментов.
b. Поведение ферментов при изменении концентрации водородных генов.
c. Высокая специфичность ферментов.
d. Ферменты не способны проникать через полупроницаемые мембраны.
e. Сохранение активности ферментами после действия водоотнимающими средствами (ацетон, спирт, нейтральные соли щелочных металлов).
Для ферментов и неорганических катализаторов характерны общие свойства:
1. Неорганические катализаторы и биологические катализаторы – ферменты требуются в небольшом количестве для проведения реакции.
2. Катализаторы биологические и неорганические выделяются после реакции в неизменном виде.
3. Катализаторы не сдвигают химического равновесия.
4. Катализаторы биологические и небиологические ведут реакцию в обход энергетического барьера, т.е. снижают энергию активации.
Отличия:
1. Ферменты обладают более высокой биологической активностью.
Пример: уреаза разлагает мочевину за 10-4 сек, а под действием воды этот процесс пройдет за 3200 лет.
Гидролиз белков
без фермента 100 – 1100С, 20% HCl – 8-10 часов
с ферментом 370С, pH 7,0 – 1-1,5 часа
7,0 – 1-1,5 часа
2. Высокая специфичность действия ферментов.
Белок 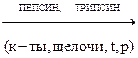 аминокислоты.
аминокислоты.
Протеолитические ферменты разрывают определенные пептидные связи. Ферменты катализируют превращение определенного субстрата или группы субстратов.
3. Активность ферментов зависит от t, pH. Ферменты действуют в более мягких условиях t=370-380, pH – организма, микроэлементы.
4. ферменты действуют последовательно и кооперативно (т.е. сообща).
S  A
A  B
B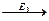 C
C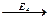 P
P
5. Локализация ферментов по отдельным компартментам.
6. Активность ферментов регулируется.
7. Скорость ферментативной реакции зависит от концентрации фермента.
Ферменты классифицируют по химической структуре:
| Ферменты (энзимы) | ||
| Простые | Сложные (холоферменты) | |
| (состоят из аминокислот, относятся пищеварительные ферменты: пепсин, трипсин, амилаза, ДНК-аза) | Белковая часть (апофермент состоит из аминокислот) | Небелковая (кофактор; витамины, нуклеотиды, ионы Ме: Fe, Mg, Co, Zn) |
Кофактор делят на 2 группы в зависимости от прочности связи:
1. кофермент – легкоотделяемая небелковая часть
2. простетическая группа – трудноотделяемая часть фермента.
Функции белковой и небелковой части фермента.
1. Белковая часть (апофермент) отвечает за активность и специфичность.
2. Небелковая часть (кофактор) отвечает за превращение субстрата («руки» фермента – разрыв связи, перенос).
Пример: Фермент аминотрансфераза состоит из белковой части и небелковой, которая представлена пиридоксальфосфатом (витамин B6). Этот кофермент обеспечивает перенос NH2-группы с аминокислоты на кетокислоту. Удаление белковой части приводит к потере специфичности (фермент начинает переносить еще 15 групп) и теряет активность.
Небелковая часть фермента неспецифична и может входить в состав разных ферментов и выполнять разные функции.
Пример: Гем входит в состав каталазы, которая обеспечивает расщепление H2O2. Гем входит также в состав цитохромов, которые осуществляют перенос электронов.
Каталаза Бел.ч Гем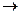 H2O2
H2O2
Цитохромы Бел.ч Гем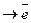
Коферменты делят:
1. Переносчики р и  - дегидрогеназы, ферменты, осуществляющие окислительно-восстановительные реакции.
- дегидрогеназы, ферменты, осуществляющие окислительно-восстановительные реакции.
Пример: НАД (витамин В5) – нуклеотид
ФАД (витамин В2) – нуклеотид
2. Переносчики различных функциональных групп – трансферазы.
Пример: Пиродоксальфосфат (ПФ) – NH2
Биотин (витамин Н) - CO2
3. Разрыв химических связей, соединение, изомеризация.
Пример: ТДФ (В1)- тиаминдифосфат
Взаимодействие фермента с субстратом идет при участии активного (ых) центров фермента.
Различают следующие виды активных центров:
1. Субстратный (якорная площадка) активный центр – обеспечивает присоединение субстрата за счет образования слабых связей: водородных, ван-дер-ваальсовых, гидрофобных взаимодействий.
2. Каталитический активный центр – отвечает за превращение субстрата. В пространстве эти центры могут быть разделены, а могут быть совмещены.
3. Аллостерический (регуляторный) обеспечивает присоединение низкомолекулярных веществ, приводит к изменению активности фермента. Аллостерический центр удален от субстратного и каталитического центров.
Закономерности построения активных центров.
1. Активные центры формируются за счет ограниченного числа аминокислот (12-16). Часто аминокислоты удалены друг от друга. Активные центры возникают при образовании четвертичной структуры.
2. В построении активных центров часто участвуют аминокислоты: гис, сер, лиз, асп, цис.
3. В построении активных центров сложных ферментов участвуют группировки кофакторов.
4. Олиго- и мультимерные ферменты на каждом протомере имеют свой каталитический и субстратный центр, аллостерический центр формируется за счет нескольких протомеров. При разрушении четвертичной структуры нарушается аллостерический центр и регуляция прекращается, а каталитическая функция характерная для протомера сохраняется.
5. Активный центр – это трехмерная структура, имеющая вид впадины или щели.
Теории, объясняющие механизм взаимодействия фермента и субстрата.
Теория Фишера – теория предшествующего соответствия, теория «ключ – замок». Согласно теории активный центр фермента существует и точно соответствует субстрату.
Недостатки (противоречия) теории:
- Нет соответствия в термодинамических расчетах (разница в расчетном количестве выделяемой энергии и практически выделяемом количестве энергии).
- По этой теории фермент может ошибаться и присоединять похожий субстрат.
- Субстраты часто низкомолекулярные вещества, а ферменты высокомолекулярные, содержащие большое число аминокислот. Теория не объясняла существование групповой специфичности.
Теория Кошленда – индуцированного соответствия, т.е. активный центр формируется в момент взаимодействия фермента и субстрата, т.е. происходит подгонка. В субстрате происходит изменение связей. Наличие активных центров определяют специфичность.
Виды специфичности.
1. Абсолбтная – одному субстрату соответствует один фермент.
Уреаза катализирует расщепление мочевины, аспартаза катализирует взаимодействие NH3 с фумаровой кислотой, в результате образуется аспарагиновая кислота, но аспартаза не действует на малеиновую кислоту.
СООН COOH
| |
CH CH2
|| 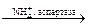 |
|
CH 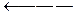 CHNH2
CHNH2
| |
COOH COOH
Фумарат Аспартат
2. Относительная групповая специфичность. Фермент расщепляет группу субстратов, для которых характерен один тип связей.
Пример: пепсин расщепляет пептидную связь –CO-NH между аминокислотами.
Аналогично действуют трипсин, химотрипсин, пептидазы.
Химотрепсин расщепляет пептидные связи между три, тир и фен, но при определенных условиях могут расщеплять амидные и сложноэфирные связи.
3. Стереохимическая специфичность – фермент обеспечивает превращение определенного оптического (стереоизомера). В организме происходит превращение L-аминокислот, но D-углеводов.
Фумараза катализирует превращение фумаровой кислоты (транс-изомер), но не действует на малеиновую кислоту (цис-изомер).
При исследовании специфичности ферментов было установленно, что молекула субстрата должна обладать двумя структурными особенностями:
1. Субстрат должен содержать специфическую химическую связь, которую фермент мог атаковать.
2. В молекуле субстрата должна быть функциональная группа, называемая связывающей группой, которая способна связываться с ферментом и ориентировать молекулу субстрата в активном центре фермента, чтобы атакуемая связь субстрата была правильно расположена по отношению к каталитической группе фермента.
Изоферменты.
Изоферменты – это множественные формы ферментов.
Изоферменты отличаются по сродству к субстрату, по максимальной скорости катализирумой реакции (по активности), по электрохимическим свойствам, по константе Михаэлиса, по тканевой локализации.
Изоферменты состоят из нескольких субъединиц.
Примером изоферментов является лактатдегидрогеназа (ЛДГ), катализующая реакцию превращения пир лак и наоборот.
ЛДГ состоит из 4 субъединиц двух разных типов – Н и М и в результате комбинаций образуется 5 изоферментов:
ЛДГ1 – Н4
ЛДГ2 – Н3М
ЛДГ3 – Н2М2
ЛДГ4 – НМ3
ЛДГ5 – М4
Изоферменты отличаются по молекулярной массе, электрофоретической подвижности, по отношению к активаторам и ингибиторам.
Для каждой ткани в норме характерно свое соотношение форм (изоферментный спектр) ЛДГ.
В сердечной мышце преобладает ЛДГ1 (Н4), а в скелетных мышцах и печени ЛДГ5 (М4).
Это свойство изоферментов используется в клинике для дифференциальной диагностики органических и функциональных поражений органов и тканей. По изменению содержания изоферментов в сыворотке крови судят о нахождении патологического процесса, так и о степени поражения органа или ткани.
| Витамин | Коферментная форма | Переносимая группа | Тип катализируемой реакции |
| В1 тиамин | ТДФ- тиаминдифосфат ТПФ (ТТФ) - тиаминпирофосфат | Отщепление СО2 | Окислительное декарбоксилирование |
| В2 рибофла-вин | ФМН – флавинмононуклеотид ФАД – флавинаденин-динуклеотид | Протоны (Н+) и электроны () | Дегидрирование |
| В3 пантоте-новая кислота | HSKoA – (аш эс коэнзим А) | Ацильная группа (R-C=O) | | Ацилирования |
| В5 никотин-амид | НАД – никотинамидаде-ниндинуклеотид НАДФ – никотинамид-адениндинуклеотид фосфат | Протоны и электроны | Дегидрирование |
| В6 пиридок-саль | ПФ – пиридоксаль фосфат | NH2-группа (амино-группа) | Трансаминирование |
| В7 биотин | Коферментной формой является сам витамин | СО2 | Карбоксилирование |
| В9,10 фолиевая кислота | ТГФК – тетрагидро-фолиевая кислота ТГФК-СН3 – метилтетра-гидрофолиевая кислота, СН2ОН–ТГФК – гидроксиметилен, ТГФК-С=О – формил- | ТГФК Н | Перенос одноуглерод-ных фрагментов: метил – СН3, гидрокси-метилен - СН2ОН, формил –С=О | H | |
| В12 кобал-амин | СН3-В12 – метил-кобаламин, ДА-В12 – дезоксиаденозил кобаламин | Перенос метильной группы СН3 | Трансметилирование |
Факторы, влияющие на активность ферментов.
- Концентрация субстрата [S]
- Концентрация фермента [F]
- Температура
- рН
- Низкомолекулярные вещества (активаторы, ингибиторы).
Концентрация субстрата зависит от питания, возраста, физической нагрузки.
Зависимость скорости ферментативной реакции субстрата выражается уравнением Михаэлиса-Ментен:
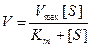
Vmax – максимальная скорость реакции
[S] – концентрация субстрата
Km – константа Михаэлиса.
Анализ уравнения Михаэлиса-Ментен.
- Концентрация субстрата мала, стремиться к нулю. При этих условиях [S] можно пренебречь:
[S]0, при этом [S] можно пренебречь:
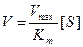
- Концентрация субстрата стремится к бесконечности, пренебрегаем Km и уравнение имеет вид:
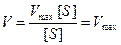
Сокращаем на [S] и скорость реакции равняется Vmax.
- Если принять, что
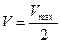 , то из уравнения Михаэлиса-Ментен, разделив его на Vmax, получили Km=[S]:
, то из уравнения Михаэлиса-Ментен, разделив его на Vmax, получили Km=[S]:
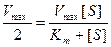 и разделив на Vmax получим
и разделив на Vmax получим 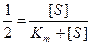 . Решая уравнение относительно Km получаем Km+[S] = 2[S],
. Решая уравнение относительно Km получаем Km+[S] = 2[S],
Km=[S]
Km – величина, численно равная концентрации субстрата при , выраженная в молях. Km = 10-1 – 10-6 – для клеток организма, величина const.
Km показывает:
- Степень сродства между ферментом и субстратом, существует обратная зависимость – чем меньше Km, тем больше сродство F к S.
- Km позволяет определить какой субстрат будет превращаться под действием данного фермента:
Например, этиленгликоль – составная часть антифриза и алкагольдегидрогеназа (АДГ) будет превращать его в щавелевую кислоту, которая является ядом для печени.
Алкагольдегидрогеназа превращает этиловый спирт в уксусный альдегид и степень сродства АДГ к С2Н5ОН выше, чем к этиленгликолю и на этом основан способ нейтрализации этиленгликоля.
- Km показывает степень сродства между белковой и небелковой частью F,
- Km позволяет определить вид ингибирования.
Способ определения Km.
- Построение график Михаэлиса-Ментен:
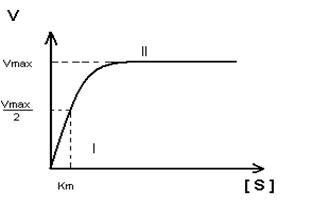
I участок – с увеличением концентрации субстрата увеличивается скорость ферментативной реакции
II участок – с увеличением концентрации субстрата скорость реакции не изменяется, т.к. все активные центры заняты.
Недостаток графика Михаэлиса-Ментен при определении Km заключается в том, что Vmax достигается с трудом, реакции в клетке протекают с оптимальной скоростью, а не Vmax.
- Построение графика Лайнуэвера-Бэрка – метод обратных величин
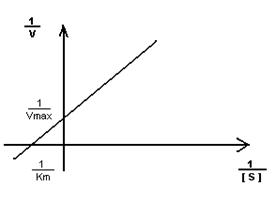
Преимущество метода заключается в том, что прямую можно построить по двум точкам и нет необходимости определять максимальную скорость.
Зависимость скорости ферментативной реакции от концентрации фермента.
Зависимость носит линейный характер. Скорость химической реакции, катализируемой данным ферментом прямопропорциональна 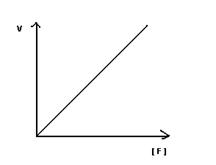 концентрации фермента. При этом концентрация субстрата величина постоянная. Это обясняетя большим количеством активных центров при определенном количестве молекул субстрата
концентрации фермента. При этом концентрация субстрата величина постоянная. Это обясняетя большим количеством активных центров при определенном количестве молекул субстрата
Зависимость скорости ферментативной реакции от температуры.
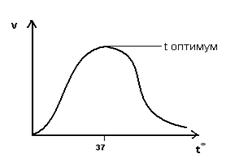 При t=36-380 ферменты обладают наибольшей активностью. Эта температура называется температурный оптимум:
При t=36-380 ферменты обладают наибольшей активностью. Эта температура называется температурный оптимум:
С повышением t0 до оптимума активность ферментов повышается.
Высокие t вызывают денатурацию ферментов.
Низкие t снижают активность ферментов.
Изменение t0 приводит к нарушению связей, закрепляющих белковую структуру ферментов (третичную, четвертичную), т.е. вызывает денатурацию.
Обратимая денатурация наблюдается при понижении t0. Это позволяет хранить ферменты, биологические жидкости, кровь.
Повышение температуры необратимо нарушает белковую структуру фермента. Это свойство используется при стерилизации материалов, инструментов.
Лихорадка – защитное свойство организма, т.к. происходит денатурация ферментов микроорганизмов и поэтому нецелесообразно применять жаропонижающие средства.

