 2015-01-21
2015-01-21 4553
4553В современной эволюционной теории вопрос о формах естественного отбора остается одним из дискуссионных. Выделяется более 30 различных форм отбора. Однако основных форм отбора в популяциях только три: стабилизирующий, движущий и дизруптивный.
Стабилизирующий отбор— форма естественного отбора, направленная на поддержание и повышение устойчивости реализации в популяции среднего, ранее сложившегося значения признака или свойства. При стабилизирующем отборе преимущество в размножении получают особи со средним выражением признака. Эта форма отбора как бы охраняет и усиливает установившуюся характеристику признака, устраняя от размножения все особи, фенотипически заметно уклоняющиеся в ту или другую сторону от сложившейся нормы (рис. 10.8).
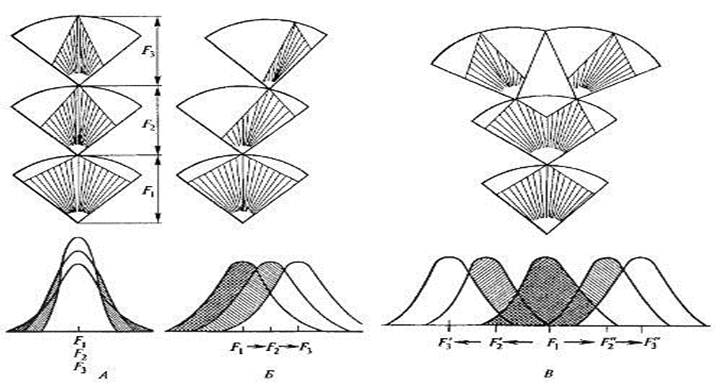
Рис. 10.8. Схема действия стабилизирующей (А), движущей (Б) и дизруптивной (В) форм естественного отбора (по Н.В. Тимофееву-Ресовскому и др., 1977). Fn — поколения. На популяционных кривых заштрихованы элиминируемые варианты. Величина дуги при отборе внутри поколения соответствует широте нормы реакции по данному признаку
Действие стабилизирующего отбора можно пояснить на многих примерах. Так, после снегопада и сильных ветров в Северной Америке было найдено 136 полуживых домовых воробьев (Passer domesticus); 72 из них выжили, а 64 погибли. У погибших птиц были очень длинные или очень короткие крылья. Особи со средними — «нормальными» — крыльями оказались более выносливыми.
Другой пример — размеры и форма цветков у энтомофильных растений. Исследование ветро- и насекомоопыляемых растений показало, что размеры и форма цветков у энтомофильных растений более стабильны, чем у анемофильных. Устойчивость цветков энтомофилов обусловлена сопряженной эволюцией растений и их опылителей, «выбраковкой» уклонившихся форм. Шмель не может проникнуть в слишком узкий венчик цветка, хоботок бабочки не может коснуться слишком коротких тычинок у растений с длинным венчиком. Только растения с цветками, строго соответствующими «стандартам», выработанным в течение миллионов лет совместной эволюцией цветковых растений и насекомых-опылителей, могут оставить потомство. Известно, что обычно наибольшее число особей в каждой популяции обладает средней выраженностью любого признака. Этот факт, подтвержденный многими тысячами исследований разнообразных признаков, служит косвенным доказательством всеобщности действия стабилизирующей формы отбора.
Один из результатов действия стабилизирующего отбора — биохимическое единство («биохимическая универсальность») жизни на Земле. Так, аминокислотный состав низших позвоночных и человека почти один и тот же, почти одинаков состав ферментов у разных растений и т. д. Биохимические основы жизни, возникшие на заре биологической эволюции, оказались надежными для воспроизведения организмов независимо от уровня их организации. Об эволюции химического состава живых организмов нередко говорят как о «теме с небольшими вариациями», что порой даже используется для доказательства отсутствия отбора вообще. На самом деле в этом проявляется действие стабилизирующего отбора.
Стабилизирующая форма отбора в течение сотен тысяч и миллионов поколений оберегает виды от существенных изменений. Она действует до тех пор, пока условия жизни, при которых данный признак или свойство выработано, существенно не меняются. Стабилизирующая форма отбора оберегает норму от разрушающего влияния мутационного процесса. Без него не было бы устойчивости в живой природе. В этом состоит важная консервативная роль естественного отбора, необходимая также для сохранения и усиления результатов движущей формы отбора.
Необходимо еще раз подчеркнуть два аспекта действия стабилизирующего отбора. С одной стороны, негативный аспект, т. е. элиминация всех особей, отклоняющихся от «стандартного» фенотипа; иногда эта форма отбора определяется как нормализующий отбор. С другой стороны, можно выделить и позитивный аспект стабилизирующего отбора, который обозначается термином канализирующий отбор (К. Уоддингтон),— отбор индивидов с генами, способными стабилизировать процесс онтогенеза и снижать его чувствительность ко всяким помехам (внутренним и внешним). В популяциях могут появляться гены, понижающие жизнеспособность фенотипа. В таких случаях отбор будет направлен в пользу особей с комплексом генов (супергенов), обеспечивающих устойчивый ход онтогенеза и способность к модификациям. Какая из двух форм стабилизирующего отбора (нормализующий или канализирующий) окажется важнее, зависит от интенсивности притока новых генов в популяцию (Э. Майр).
Движущий отбор.Движущей (или направленной) формой отбора принято называть отбор, способствующий сдвигу среднего значения признака или свойства. Такой отбор способствует закреплению новой нормы взамен старой, пришедшей в несоответствие с условиями. Изменение признака при этом может происходить как в сторону усиления, так и ослабления. Большинство приводившихся ранее прямых и косвенных доказательств естественного отбора относится именно к движущей форме, отражающей классическое, дарвиновское, понимание сущности отбора.
Утрата признака — обычно результат действия движущей формы отбора. Например, в условиях функциональной непригодности органа (или его части) естественный отбор способствует их редукции. Утрата крыльев у части птиц и насекомых, пальцев у копытных, конечностей у змей, глаз у пещерных животных, корней и листьев у растений-паразитов — примеры действия движущего отбора в направлении редукции органов. Материал для действия отбора в направлении редукции органов доставляется разного рода мутациями, которые ведут к дезинтеграции организма и нарушению системы его корреляций (см. гл. 14).
Была экспериментально изучена возможность изменения числа щетинок у дрозофилы (Drosophila melanogaster). Направленный отбор через 30 поколений в одной линии снизил их число с 32 до 25, а в другой (после 20 поколений) повысил до 45—50 шт. (см. рис. 9.1). Генетический анализ мух новых линий показал наследственную обусловленность вновь возникших особенностей. В экспериментах с тем же видом дрозофилы показана возможность заметного повышения холодоустойчивости после отбора. Так, у исходной линии при температуре 0°C выживало лишь 28% четырехдневных личинок. После 30 поколений отбора для личинок температура 0"С превратилась в фактор, стимулирующий развитие. Этот процесс характерен в природе; часто первоначально вредный фактор при его постоянном действии превращается в фактор нормального развития. Анализ показал, что холодоустойчивость в эксперименте возникла не в результате распространения единичной мутации, вызванной действием холода, а в результате действия движущего отбора в пределах гетерогенной популяции.
Дизруптивный отбор.Эта форма отбора существует в тех случаях, когда ни одна из групп генотипов не получает абсолютного преимущества в борьбе за существование из-за разнообразия условий, одновременно встречающихся на одной территории. При этом в одних условиях отбирается одно качество признака, в других — другое. Дизруптивный отбор направлен против особей со средним и промежуточным характером признаков и ведет к установлению полиморфизма в пределах популяций. Популяция как бы «разрывается» по данному признаку на несколько групп. Поэтому дизруптивный отбор называется также разрывающим или расчленяющим. Итак, дизруптивной называется форма отбора, благоприятствующая более чем одному фенотипу и действующая против средних промежуточных форм.
Во всех случаях полиморфизма действовал отбор дизруптивной формы. Рассмотрим пример действия такого отбора по закреплению полиморфизма по окраске раковин у земляной улитки (Cepaea nemoralis). Эти улитки различаются по количеству полос на раковине и степени пигментированности входного отверстия раковины в зависимости от окраски фона. Так, в лесах, где почвы коричневого цвета, чаще встречаются особи с коричневой и розовой окраской раковин, на участках с грубой и желтой травой преобладает желтая окраска и т. п. Подобные различия в окраске раковин явно приспособительны, так как предохраняют животных от истребления птицами. Полиморфизм улиток по окраске — результат действия дизруптивного отбора против средней нормы: единая популяция распадается на несколько форм и ни одна из них не имеет решающего преимущества перед другой.
Общий результат дизруптивного отбора — формирование гетерозиготного полиморфизма. Результат дизруптивного отбора (и возникновение его) можно представить в виде совместного и попеременного действия ведущего и стабилизирующего отбора.
Другие формы естественного отбора.Как уже говорилось, к настоящему времени выделено несколько десятков разных форм естественного отбора, связанных с отдельными специфическими сторонами действия единого, всеобщего («дарвиновского») естественного отбора. Рассмотрим некоторые из них.
Часто оказывается так, что определенный генотип имеет селективное преимущество только тогда, когда он редок в популяции, и немедленно теряет это преимущество, оказавшись в избытке. Это типичная ситуация действия частотно-зависимого отбора. Именно под его контролем находятся многие признаки покровительственной окраски типа мимикрии (например, при апостатическом отборе — одной из форм частотно-зависимого отбора — полиморфизм в популяции жертвы может поддерживаться избирательным хищничеством, направленным на более многочисленную форму).
Когда в лабораторной популяции дрозофил менялось соотношение самцов, ведущих свое происхождение из двух разных природных популяций, самцы, оказавшиеся в меньшинстве, участвовали в спариваниях непропорционально чаще самцов, составляющих большинство. Во всех подобных случаях приспособленность генотипа (т. е. положительное давление отбора) будет функцией частоты этого генотипа.
Другая большая группа примеров действия естественного отбора связана с выбором одной из двух главных стратегий размножения: либо короткая продолжительность жизни, ранняя половая зрелость и огромное число потомков, либо длительная жизнь особи, позднее наступление зрелости, небольшое число потомков. И в первом (r-стратегия и соответственно r-отбор), и во втором случае (K-стратегия и соответственно K-отбор) достигается в конечном итоге сохранение популяции. При r-отборе большая часть энергетических затрат вида идет на появление сравнительно многочисленного потомства (обычно в нестабильных условиях), при K-отборе — на выживание исходно немногочисленного потомства (обычно в стабильных условиях среды). Рассмотренные формы отбора могут быть названы в общей форме плотностно-зависимым отбором (т. е. отбором, связанным с плотностью населения популяции).
Из других более частных форм отбора упомянем дестабилизирующий отбор (Д.К. Беляев) и отбор местообитания. Под дестабилизирующим отбором имеется в виду разрушение коррелятивных связей в организме при интенсивном отборе в каком-то определенном направлении. Так, например, отбор на уменьшение агрессивности у хищных зверей в неволе ведет к дестабилизации цикла размножения (животные могут размножаться круглый год). Последняя форма отбора может быть проиллюстрирована таким примером: самки двух близких видов дрозофил Гавайских островов откладывают яйца на одном и том же виде растений, но самки одного вида откладывают яички на теневой стороне растения, тогда как другого — исключительно на солнечной стороне. Отбор местообитаний определяет более полное использование пространства близкими симпатрическими видами (ведет к увеличению суммы жизни, как образно заметил в свое время Ч. Дарвин, не выделявший никаких специальных форм естественного отбора, но в своих работах прозорливо описавший практически все основные формы действия отбора).








